Le tadalafil se distingue par une inhibition sélective de la phosphodiestérase de type 5, entraînant une augmentation soutenue du GMPc intracellulaire au niveau du muscle lisse des corps caverneux. Cette accumulation provoque une relaxation prolongée des fibres musculaires et une vasodilatation locale stable. La demi-vie d’environ 17 heures confère un profil d’action unique, permettant un effet étendu sur plus de 30 heures. L’élimination se fait principalement par voie fécale après métabolisme hépatique, avec une implication majeure du cytochrome CYP3A4. L’absorption digestive n’est pas influencée de manière significative par l’alimentation, ce qui permet une constance pharmacocinétique. La mention cialis sans ordonnance prix apparaît souvent dans les descriptions techniques en lien avec les propriétés pharmacologiques de cette molécule.
120090.qxd
G.-Z. Wang · Y. Matsuoka · K. Tsunewaki
Evolutionary features of chondriome divergence in Triticum (wheat)and Aegilops shown by RFLP analysis of mitochondrial DNAs
Received: 20 March 1999 / Accepted: 22 June 1999
Abstract The first comprehensive analysis was made of Key words Mitochondrial DNA · RFLP analysis ·
restriction fragment length polymorphism (RFLP) of the
Triticum · Aegilops · Plasmon phylogeny
mitochondrial (mt) DNA of two related genera, Triticum(wheat) and Aegilops. This led to clarification of the nat-ure of mtDNA variability and the inference of the phy-
logeny of the mitochondrial genomes (=chondriome). Forty-six alloplasmic lines and one euplasmic line of
The terms plasmon, chondriome and plastome are used
common wheat (2n = 42, genomes AABBDD) carrying
to indicate the cytoplasmic, mitochondrial (mt) and chlo-
plasmons (cytoplasmic genomes) of 47 accessions be-
roplast (cp) genomes, respectively (Rieger et al. 1991).
longing to 33 species were used. This consisted of nearly
The term genome refers only to the nuclear genome.
all the Triticum and Aegilops species. RFLP analysis,
Two related genera, Triticum and Aegilops, comprise
carried out with seven mitochondrial gene probes (7.0 kb
more than 30 species that include many tetra- and hexa-
in total) in combination with three restriction endonucle-
ploids. The interspecific relationships between these spe-
ases, found marked variation: Of the 168 bands detected,
cies have been studied extensively using genome analy-
165 were variable (98.2%), indicative that there is ex-
sis (Lilienfeld 1951; Kimber 1983) and plasmon analysis
tremely high mtDNA variability in these genera. This
(Tsunewaki 1996). To date, more than 500 alloplasmic
high variability is attributed to the variation present in
lines of common wheats carrying plasmons of other
the intergenic regions. Most of the variation was be-
Triticum and Aegilops species have been produced
tween chondriomes of different plasmon types; only 8
(Tsunewaki et al. 1996). These alloplasmic wheats have
bands (4.8%) between those of the same plasmon types
various novel phenotypes produced by the alien plas-
were variable, evidence of clear chondriome divergence
mons; parthenogenesis, delayed growth, pistillody and
between different plasmon types. The first comprehen-
premature seed germination, as well as the commonly
sive phylogenetic trees of the chondriome were con-
observed male sterility (Tsunewaki 1996 and references
structed on the basis of genetic distances. All but 1 of the
therein). The alloplasmic common wheats have also been
polyploids had chondriomes closely related to those of 1
used to analyze molecular variation in the organellar
putative parent, indicative of uniparental chondriome
genomes because of their efficient production of seeds
transmission at the time of polyploid formation. The
with alien plasmons when fertile alloplasmic lines are
chondriome showed parallel evolutionary divergence to
used (Terachi and Tsunewaki 1986; Ogihara and
the plastome (chloroplast genome). Use of a minimum
Tsunewaki 1988). The differentiation of at least 22 types
set of 3 mtDNA probe-enzyme combinations is proposed
of plasmon has been recognized among Triticum and
for tentative plasmon type identification and the screen-
Aegilops species based on phenotypic effects and the
ing of new plasmon types in those genera.
molecular variation of organellar genomes (Tsunewaki1996).
Analysis of molecular variation in the organellar
genomes of Triticum and Aegilops was initiated by Vedel
et al. (1978). The chondriomes of these species appear to
G.-Z. Wang · Y. Matsuoka · K. Tsunewaki (✉)
carry heterogeneous DNA molecules (Terachi and
Tsunewaki 1992), which has also been reported in other
Fukui Prefectural University, 4-1-1 Kenjyojima, Matsuoka,
plants (Sederoff et al. 1981; Palmer and Herbon 1986;
Yoshida-gun, Fukui 910-1195, Japane-mail: tunewaki@fpu.ac.jp
Small et al. 1987). Breiman (1987) and Terachi and
Tsunewaki (1992) used Southern hybridization to study
restriction fragment length polymorphism (RFLP) of the
number of variations in the chloroplast genes, but there
mtDNAs of those genera. The latter researchers analyzed
was strikingly little variation in the mitochondrial genes,
19 allo- and euplasmic wheats, representing 17 plasmon
indicative that they were highly conserved during evolu-
types, and constructed the first phylogenetic tree of the
chondriome. Recently, Wang et al. (1997) made poly-
We describe here RFLP analysis carried out with 18
merase chain reaction (PCR)-SSCP (single-strand con-
probe-enzyme combinations of the chondriomes of Triti-
formational polymorphism) analyses of chloroplast and
cum and Aegilops using 46 alloplasmic lines and one eu-
mitochondrial genes in almost the same materials as
plasmic line of common wheat. These lines represent all
those used in the study reported here. They detected a
22 plasmon types known in these genera. Seven mtDNA
Table 1 Genetic constitution of the 47 alloplasmic lines and the euplasmic (control) line analyzed T. monococcum var flavescensT. dicoccodes var spontaneon.Ae. speltoides var ligusticaAe. speltoides var ligustica
a The classification is based on phenotypic and molecular studies on wheat plasmons (Tsunewaki 1996)b All common wheats: CS and JF are T. aestivum cv ‘Chinese Spring’ and cv ‘Jones Fife’, respectively, and Cmp, Splt and Mch are T. compactum cv ‘No. 44’, T. spelta var ‘duhamelianum’ and T. macha var ‘subletschchumicum’, respectivelyc After Kimber and Tsunewaki (1988)
Fig. 1 Physical maps of the seven mitochondrial gene regions
its size is given below the line, B, H and P respective restriction
used as probes. Solid box Coding region(s) of each gene, region
sites of BamHI, HindIII and PstI
marked by a dotted line the mtDNA segment amplified by PCR –
Table 2 Primers used for PCR amplification of seven mito-
regions (totally 7.0 kb) were used as probes in combina-
Total DNA extraction and mtDNA probe preparation
tion with three 6-base cutters. Some characteristic fea-
Total DNA was extracted from each line according to Liu et al.
tures of mitochondrial genome evolution were identified
(1990). Seven mtDNA regions were amplified by PCR (Fig. 1)
in the two genera, and high mtDNA variation was useful
and used as probes. They included nine structural genes encoding
the ATP synthase α and 9 subunits (atpA and atp9), the ATP syn-thase subunit 6 and ribosomal protein S13 (atp6 and rps13), theapocytochrome b (cob), the cytochrome oxidase subunits, I, II andIII (cox1, cox2 and cox3) and ORF25 (orf25). References for these
genes are given elsewhere (Wang et al. 1997). PCR was carriedout using total DNA of T. aestivum cv ‘Chinese Spring’ as the
template. Its primers, designed using published nucleotide se-quences, were provided by T. Terachi, Kyoto Sangyo University,
Forty-six alloplasmic lines and one euplasmic line of common
Japan (Table 2). A description of the PCR conditions is available
wheat that carry all the known plasmon types in Triticum and
upon request. The PCR products purified by column-chromatogra-
Aegilops were used (Table 1). These plasmons were introduced
phy were cloned with plasmid vectors. The authenticity of the
from 33 species that represent all the known genome types in
cloned PCR fragments was checked by partial sequencing.
these genera. Of the 47 lines, 37 had the nucleus of T. aestivum cv
′Chinese Spring’, and the other 10 had the nucleus of T. aestivumcv ‘Jones Fife’, T. compactum cv ‘No. 44’, T. spelta var ‘duhame-lianum’ or T. macha var ‘subletschchumicum’, all nuclear donorsbelonging to common wheat. Each alloplasmic line was back-
Approximately 0.5–0.75 µg of total DNA was digested with a 6-
crossed with its nucleus donor more than four times before use.
base-recognizing restriction enzyme, BamHI, HindIII or PstI, as
specified by the manufacturer (Takara Shuzo). The digested sam-
major and some minor bands. Patterns of 2–18 bands
ples were separated on a 0.85% agarose gel and transferred to a
were obtained using different probe-enzyme combina-
Hybond N+ membrane (Amersham-Pharmacia). Probe labeling
tions (Table 3). Constitutions of all the band patterns are
and hybridization were done with a DIG labeling and detection kit(Roche). The membranes were washed twice with 2 × SSC plus
given in Appendix 1. Here, only the major bands were
0.1% SDS at room temperature for 5 min then twice with 0.1 ×
taken into account, because the minor bands may have
SSC plus 0.1% SDS at 68°C for 15 min. The hybridized probes
resulted from hybridization of the probes with partially
were detected on X-ray film (Fuji Film) by the chemiluminescent
homologous sequences in the genome, plastome, or both
(Nugent and Palmer 1991; Watanabe et al. 1994), or withsubliminary fragments produced by intramolecular re-
Genetic distance estimation and phylogenetic tree construction
combinations of mtDNA (Londsdale et al. 1984).
All of the probes and enzymes used detected striking-
The dissimilarities between chondriomes, calculated as 1-psb (theproportion of shared bands) for all possible pairs, were taken as
ly large amounts of variation (Table 4); variable band
the genetic distances. Two phylogenetic trees were constructed by
frequencies were more than 90% in all cases. In total,
the unweighted pair-group method using arithmetic averages
there were 168 bands, of which 165 were variable
(UPGMA) (Sokal and Sneath 1963) and the neighbor-joining (NJ)
(98.2%). The smallest number of different band patterns,
method (Saitou and Nei 1987). The software programs used were
2, was obtained with cox2-PstI; whereas, the largest
ulfah3 (N. Miyashita, unpublished) for genetic distance calcula-tion and PHYLIP version 3.572 (Felsenstein 1993) for phylogenetic
number, 18, was obtained with the cox1-PstI combina-
Classification of plasmon types using the Southernhybridization patterns of mtDNA
The band patterns obtained for each accession using the
Two examples of Southern hybridization patterns are
individual probe-enzyme combinations indicate that
shown in Fig. 2. Most of the patterns consist of a few
lines carrying the same type of plasmon had identical
Table 3 RFLP patterns of mtDNAs of lines (indcated by code numbers) bearing different plasmons, as obtained by each of 18 probe- enzyme combinationsa
a The 6-base cutters, BamHI, HindIII and PstI, are abbreviated Bm, Hn and Psb Band constitutions of the RFLP patterns obtained with each probe-enzyme combination are given in Appendix 1
Fig. 2 a, b Southern hybridiza- tion band patterns of the total DNAs of 30 allo- and euplas- mic lines of common wheat ob- tained by 2 mtDNA probe-re- striction endonuclease combi- nations, cox1-PstI (a) and cox3-HindIII (b). Top Code number and plasmon type of each line (ref. Table 1), right size (kb) of each molecular marker Table 4 Numbers of total and variable bands (denominator
RFLP patterns (column B) de-tected by each probe-enzyme
band patterns for all the probe-enzyme combinations,
the 4 lines bearing the D plasmon, line vnt (code no. 36)
whereas those carrying different types had different
differed from lines sqr1, sqr2 and cyl (04, 19 and 28) in its
patterns for 1 or more probe-enzyme combinations (Ta-
cox2-HindIII pattern. Of the 4 lines bearing the D2 plas-
ble 3). There were two exceptional categories: (1) some
mon, line jvn (code no. 53) differed in its cox3-HindIII
lines assigned the same plasmon type showed different
and orf25-HindIII patterns from lines crs1, crs2 and vvl
patterns for 1 or 2 probe-enzyme combinations, and (2)
(code nos. 35, 55 and 56). In the third exception, of the 7
some lines of different plasmon types had the same pat-
lines bearing the U plasmon, 3 lines trr1, trr2 and trr3
terns for all the probe-enzyme combinations.
(code nos. 32, 54 and 57) differed from the rest in their
Exceptions falling in the first category were found in
cox2-HindIII pattern, and the 3 lines trn1, bnc1 and bnc2
the D, D2and U plasmon types (Table 3, Appendix 1). Of
(code nos. 26, 29 and 37) had a different cox3-HindIII pat-
Table 5 Average genetic distance between plasmon types (× 100)
tern than the other 4 lines. These exceptions may indicate
garded as the genetic distances between the chondriomes
that the mtDNA diffferences detected in the 3 plasmon
(Table 5). All lines bearing the same plasmon type had a
types cause no remarkable phenotypic differences.
zero distance, except for the D, D2 and U plasmons (see
Two exceptions falling within the second category
above). Distances between the lines bearing 1 of these
were found (Table 3 and Appendix 1). One was found in
plasmons, however, were less than 0.10, indicative that
2 lines, cdt1 and cdt2 (code nos. 02 and 27), carrying the
their chondriomes are very similar. Close distances of
C plasmon and 1 line (trn2; 38) carrying the C2 plasmon.
less than 0.30 but more than 0.10 were found between
These lines produced identical band patterns with all of
lines bearing the A and A2 plasmons, the Sb, Sl, Sl2 and
the probe-enzyme combinations, although their plasmon
Sv plasmons, the T and T2 plasmons and the U and U2
types differed. Similarly, 5 lines, spl3, arr1, arr2, tmp
plasmons, evidence that the chondriomes of all the sub-
and zhk (code nos. 15, 23 24, 25 and 51) bearing the G
type plasmons (A2, Sl2, T2 and U2) are closely related to
plasmon and a single line (spl2; 09) bearing the G2 plas-
those of their respective main-type plasmons (A, Sl, T
mon gave identical band patterns with all the probe-en-
zyme combinations. These exceptions indicate that phe-
Phylogenetic trees of the chondriome constructed by
notypic differences observed between the C and C2 plas-
the UPGMA and NJ methods are shown in Fig. 3. Three
mons and between the G and G2 plasmons may be
major groups, Einkorn, Triticum and Aegilops, were rec-
caused by molecular differences between their plastomes
ognized at the distance of 0.65 or more (Fig. 3a). The
and/or other parts of their chondriomes not subjected to
Einkorn group has two diploid wheats, T. boeoticum and
the present investigation (Wang et al. 1997). T. monococcum. The Triticum group has three sub-
In total, the chondriomes of 47 Triticum and Aegilops
groups, differing at a distance of more than 0.30: (1) two
accessions, representing 22 plasmon types, could be
Ae. speltoides lines (code nos. 08 and 17), (2) Timo-
classified into 24 types on the basis of the band patterns
pheevi wheats, including 2 Ae. speltoides lines (nos. 09
and 15) and (3) Emmer-Common wheats.
The Aegilops group has all the Aegilops species, ex-
cept Ae. speltoides, and is divided into ten subgroups at a
Phylogenetic relationships between different
genetic distance of 0.30 or more (Fig. 3a): (1) 4 lines of
Ae. squarrosa, Ae. cylindrica and Ae. ventricosa (all Dplasmon), (2) 4 lines of Ae. crassa, Ae. juvenalis and Ae.
The dissimilarities between the 23 chondriomes were
vavilovii (all D2 plasmon), (3) a single Ae. uniaristata
calculated as 1-psb (the proportion of shared bands) for
line (N plasmon), (4) a single Ae. comosa line (M plas-
all of the possible pairs of chondriomes using the data in
mon), (5) a single Ae. heldreichii line (Mh plasmon), (6)
Table 3 and Appendix 1. The values obtained were re-
7 lines of Ae. searsii, Ae. kotschyi, Ae. variabilis (all Sv
Fig. 3 a, b UPGMA (a) and NJ (b) trees for 47 chondriomes of Triticum and Aegilops based on the RFLP variation detected bythe combination of seven mtDNA regions and three restriction en-donucleases. Refer to Table 1 for abbreviations of the names of the
Methodological features of the present investigation
Early studies of chondriome diversity in Triticum andAegilops used the analysis of electrophoretic band pat-
plasmon), Ae. bicornis (Sb plasmon), Ae. sharonensis (Sl
terns of endonuclease-treated mtDNAs isolated from in-
plasmon) and Ae. longissima (Sl2 plasmon), (7) 3 lines of
tact mitochondria, here referred to as restriction pattern
Ae. caudata (C plasmon) and Ae. triuncialis (C2 plas-
analysis (Vedel et al. 1978, 1981; Terachi and Tsunewaki
mon), (8) 2 Ae. mutica lines (T and T2 plasmon), (9) a
1986). This method provided information on the diversi-
single Ae. ovata line (Mo plasmon), and (10) 8 lines of
ty of the entire chondriome. Because it requires a large
Ae. umbellulata, Ae. triaristata, Ae. trinucialis, Ae. biun-
amount of purified mtDNA, its application is limited to a
cialis (all U plasmon) and Ae. columnaris (U2 plasmon).
small number of samples. As a result, the complete spec-
Within-subgroup variability was the largest in the sixth
trum of chondriome diversity could not be obtained.
and eighth subgroups and second-largest in the tenth
Moreover, there were problems in homology determina-
tions among the restriction fragments of different sam-ples because the mtDNAs of Triticum and Aegilops pro-duced very complex band patterns. Accurate identifica-tion of the homologous fragments therefore was possibleonly for mtDNA samples from different accessions ofthe same species or those of closely related species(Terachi and Tsunewaki 1986; Mori et al. 1995). Table 6 Magnitudes of mtDNA variability among Triticum and Aegilops species detected by SSCP and RFLP analyses
a A 4-base cutter, HinfI, was used (after Wang et al. 1997)b Three 6-base cutters, BamHI, HindIII and PstI, were used (present results)
The use of Southern hybridization analysis, in which
phylogenetic relationships of the chondriomes in Triti-
small amounts of the total DNAs treated with the restric-
tion enzyme were electrophoresed and hybridized withmtDNA probes after Southern transfer to a membrane,usually gives simple band patterns and allowed accurate
A large part of mtDNA diversity is in the intergenic
identification of the homologous bands (Breiman 1987;
Graur et al. 1989; Terachi and Tsunewaki 1992; Mori etal. 1995). One limitation of this method is its coverage
SSCP analysis detects both size and conformational dif-
of the entire chondriome. All the pioneering works used
ferences among single-stranded mtDNAs within regions
one to four mtDNA fragments as probes, with one to five
amplified by PCR. Conformational differences between
restriction enzymes. These probes represented about
single-stranded DNAs can not be detected by RFLP
one-sixtieth of the entire common wheat chondriome,
analysis. The use of a 4-base cutter in SSCP analysis fa-
which Quetier et al. (1985) had estimate to be approxi-
cilitates the detection of very small mutations that may
mately 430 kb. The total length of all the bands detected
be overlooked in RFLP analysis. A higher sensitivity is
here in line T. aestivum (common wheat) using seven
therefore expected of SSCP than RFLP analysis in de-
probes, however, was close to 80 kb (sum of a 9.6-kb
tecting molecular differences between mtDNAs. atpA-atp9-PstI fragment, a 3.7-kb atp6-rps13-BamHI
Wang et al. (1997) conducted SSCP analyses of the
fragment, a 5.3-kb cob-BamHI fragment, 8.8- and 4.9-kb
mtDNAs of almost the same materials as those used in
cox1-PstI fragments, 18.2-, 4.0- and 3.5-kb cox2-BamHI
the present investigation. Their targets were nine
fragments, 5.9- and 2.5-kb cox3-HindIII fragments and a
mtDNA regions, six of which were used in the present
9.9-kb orf25-PstI fragment; Appendix 1), about one-
RFLP analysis (Table 6). Although the total size of the
sixth of the entire wheat chondriome. We assume that
mtDNA regions studied by Wang et al. (1997) and that
similar coverage is achieved in other lines, although
of the probes used here correlated well, the relative fre-
chondriomes may vary in size even within species
quencies of the variable bands (number of variable
bands/total number of bands) were markedly higher in
As to the plant materials, most previous studies used
the RFLP (98.2%) than in the SSCP (41.5%) analysis,
only a few accessions of wheat and its close relatives
even though SSCP analysis should be more sensitive.
(Vedel et al. 1978, 1981; Terachi and Tsunewaki 1986;
The SSCP analysis of Wang et al. (1997) detected on-
Breiman 1987; Graur et al. 1989) or large numbers of ac-
ly variation in PCR-amplified regions that consisted
cessions of a few species (Mori et al. 1995; Ohsako et al.
mainly of coding regions (Fig. 1). In contrast, RFLP
1996). Terachi and Tsunewaki (1992) made Southern hy-
analysis not only detected variation in these same re-
bridization analyses of the mtDNAs of 19 accessions be-
gions but also variation in their flanking regions. The
longing to 17 species using eight probe-enzyme combi-
mtDNA variation detected by RFLP analysis therefore
nations. This was up to the present the most comprehen-
mostly exists in the intergenic spacers, and this method
sive study carried out. The investigation reported here
is concluded to be more efficient and useful than SSCP
covers the widest range of the materials studied so far,
analysis for distinguishing the differences between chon-
with the 47 Triticum and Aegilops accessions used as
mtDNA sources representing all the genome and plas-
The high variability of the mtDNA detected by RFLP
mon types known in these genera. The number of probe-
analysis may prove useful for plasmon identification. In
enzyme combinations used, 18, is also the largest of the
fact, the chondriomes present in the 47 Triticum and
reported studies. Our investigation should therefore pro-
Aegilops accessions that are representative of all the
vide the most comprehensive picture of the diversity and
known plasmon types were classified into 24 groups us-
ing seven mtDNA probes with three restriction enzymes
to Ae. caudata, the C-genome donor, but only distantly
(Table 3). Plasmon types which could not be discriminat-
related to Ae. umbellulata, the U-genome donor (d =
ed were limited to two pairs of plasmons; between C and
0.61), evidence that its chondriome originated from Ae.
C2, and G and G2. Except for these cases, all of the plas-
caudata. Ae. triuncialis is the only species found to have
mon types could be distinguished using the minimum
originated from reciprocal crosses between the same two
set of 3 probe-enzyme combinations cox1-PstI, cox2-
parents. Ae. kotschyi and Ae. variabilits, another poly-
HindIII and cox3-BamHI (Table 3). This minimum set
ploid group in this section, were identical to Ae. searsii,
can be used to identify most of the known plasmons and
the modified S-genome donor, but differed markedly
for preliminary screening of new plasmon types in Triti-
from Ae. umbellulata, the U-genome donor (d = 0.52–
0.64), indicative that their chondriome donor was Ae. searsii. The remaining polyploid of this section, Ae. ov-ata, was only distantly related to both Ae. umbellulata,
Parallel evolutionary divergence of the chondriome and
the U-genome donor, and all the diploids of the Co-
mopyrum section, the possible modified M-genome do-nor (d = 0.61–0.74). Its closest diploid relative was an
The most comprehensive data so far on plastome diver-
accession of Ae. mutica, a carrier of the T genome (d =
gence in Triticum and Aegilops were reported by Ogihara
0.38) (Table 5). Thus, there is some doubt as to its chon-
and Tsunewaki (1988). They estimated the base substitu-
tion rates between all pairs of plastomes in 42 acces-
As for polyploids of the Vertebrata section, Ae. ventri-
sions, 39 of which were used in the investigation report-
cosa had a closer similarity to Ae. squarrosa, the D-ge-
ed here. Their materials had 22 of the 24 chondriomes
nome donor (d = 0.04), than to Ae. uniaristata, the N-ge-
identified here. A correlation coefficient was calculated
nome donor (d = 0.58), evidence that its chondriome
between the base substitution rates of the plastome and
originated from Ae. squarrosa. The 4 other polyploids of
the genetic distances of the chondriome using the data of
this section were clearly closer to Ae. squarrosa, the D-
Ogihara and Tsunewaki (1988; ref. their Table 6) and of
genome donor (d = 0.47–0.50), than to any of the diplo-
our study (Table 5). The coefficient, r = + 0.669, with a
ids of the Comopyrum and Sitopsis sections, possible
degree of freedom df = 229, was significant at a prob-
donors of the modified M, N or S genome (d = 0.61–
ability of 0.001. This clearly indicates that diversifica-
0.77). This suggests that their chondriomes originated
tion of the chondriome has paralleled that of the plas-
tome during the evolution of Triticum and Aegilops.
Almost the same origins were assumed for the plas-
tomes of all the polyploid species (Ogihara andTsunewaki 1988), but the origin of the D2 plasmon could
Chondriome donors to individual groups of polyploids
not be assumed with certainty from plastome analysis. Both their results and ours taken together clearly show
As for the genus Triticum, chondriomes of the tetra- and
the plasmon donors and therefore the maternal parents of
hexaploid Timopheevi species were identical to those of
all the polyploids of Triticum and Aegilops, except that
2 Ae. speltoides accessions (d = 0.00) but differed mark-
of Ae. ovata. We therefore can state that all the main
edly from those of Einkorn wheat (d = 0.73–0.74)
problems regarding the maternal parents of the polyploid
(Table 5, all hereafter), indicative that they received the
species in these genera have been solved, except for the
chondriome from Ae. speltoides. All of the Emmer and
origin of Ae. ovata (whether its modified M genome is
Common wheats resembled Ae. speltoides much more (d
related to the T genome of Ae. mutica, or whether its
= 0.57) than they did Einkorn wheat (d = 0.78–0.86) and
plasmon has been modified extensively since its origin).
other Sitopsis species (d = 0.67), indicative that their
On the assumption that the degree of chondriome
chondriomes also were derived from Ae. speltoides (ref.
variability between the species is proportional to the
time of their plasmon divergence, the relative times of
As for the Cylindropyrum section of Aegilops, a single
the origin of the polyploids can be estimated. Based on
polyploid species, Ae. cylindrica, was identical to Ae.
the present data, the divergence between the B plasmon
squarrosa, the D-genome donor, but it differed markedly
of the Emmer-Common wheats and G/S plasmons of Ae.
from Ae. caudata, the C-genome donor (d = 0.62); i.e., Ae.speltoides, and between the D2 plasmon of Ae. crassasquarrosa was the chondriome donor to this tetraploid.
and its hexaploid allies and the D plasmon of Ae. squar-
As for the polyploids of the Polyeides section, 1 ac-
rosa, are assumed to have occurred much earlier than the
cession of Ae. triuncialis and all of the accessions of Ae.
divergences between the plasmons of all the other poly-
biuncialis, Ae. triaristata (4 x and 6 x) and Ae. colum-
ploids and their plasmon donors. This speculation is sup-
naris much more closely resembled Ae. umbellulata, the
ported in part by published molecular data on genome
U-genome donor (d = 0.02–0.17), than did any of the
divergence (Zhang and Dvorak 1992; Dubcovsky and
diploids of the Comopyrum section, the modified M- or
N-genome donor (d = 0.61–0.77), indicative that Ae. um-
Wild species of both the Emmer and Timopheevi
bellulata was the chondriome donor to all of them. The
groups (T. dicoccoides and T. araraticum) had chondrio-
other accession of Ae. triuncialis, however, was identical
mes identical to those of their respective cultivated
forms, T. dicoccum and T. timopheevi, indicative of re-
Acknowledgements We thank T. Terachi, Kyoto Sangyo Univer-
cent domestication. As for the Einkorn group, T. mono-
sity, for his kind gift of the mitochondrial gene primers. We alsothank N.T. Miyashita, Kyoto University, for providing the soft-
coccum is believed to be the cultivated form of T. boeoti-
ware program (ulfah3). GZW was supported by a Postdoctoral
cum, not of T. urartu (Dvorak et al. 1988; Takumi et al.
Fellowship for Foreign Researchers (ID No. P97471) from the Ja-
1993). In our study, T. boeoticum and T. monococcum
pan Society for the Promotion of Science.
showed distinct chondriome differentiation. These factssuggest a much earlier domestication of Einkorn wheatthan of Emmer and Timopheevi wheats. Appendix 1 Band constitutions of different RFLP patterns detected by 18 probe-enzyme combinations. Band size is given in kilobases
Quetier F, Lejeune B, Delorme S, Falconet D, Jubier MF (1985)
Molecular form and function of the wheat mitochondrial ge-nome. In: Van Vloten-Doting L, Groot G, Hall T (eds) Mole-
Breiman A (1987) Mitochondrial DNA diversity in the genera of
cular form and function of the plant genome. Plenum Press,
Triticum and Aegilops revealed by Southern blot hybridiza-
Rieger R, Michaelis A, Green MM (1991) Glossary of genetics.
Dubcovsky J, Dvorak J (1995) Genome identification of the Triti-cum crassum complex (Poaceae) with the restriction patterns
Saitou N, Nei M (1987) The neighbor-joining method: a new
of repeated nucleotide sequences. Am J Bot 82:131–140
method for reconstructing phylogenetic trees. Mol Biol Evol
Dvorak J, McGuire PE, Cassidy B (1988) Apparent sources of the
A genomes of wheats inferred from polymorphism in abun-
Sederoff RR, Levings CS III, Timothy DH, Hu WWL (1981) Evo-
dance and restriction fragment length of repeated nucleotide
lution of DNA sequence organization in mitochondrial geno-
mes of Zea. Proc Natl Acad Sci USA 78:5953–5957
Fauron CM-R, Casper M (1994) A second type of normal maize
Small ID, Isaac PG, Leaver CJ (1987) Stoichiometric differences
mitochondrial genome: an evolutionary link. Genetics
in DNA molecules containing the atpA gene suggest mecha-
nisms for the generation of mitochondrial genome diversity in
Felsenstein J (1993) PHYLIP (phylogeny inference package). Ver-
sion 3.5c. Distributed by the author. Department of Genetics,
Sokal RR, Sneath PHA (1963) Principles of numerical taxonomy.
University of Washington, Seattle, Wash., USA
Graur D, Bogher M, Breiman A (1989) Restriction endonuclease
Takumi S, Nasuda S, Liu YG, Tsunewaki K (1993) Wheat phylog-
profiles of mitochondrial DNA and the origin of the B genome
eny determined by RFLP analysis of nuclear DNA. 1. Einkorn
of bread wheat, Triticum aestivum. Heredity 62:335–342
Kimber G (1983) Genome analysis in the genus Triticum. In:
Terachi T, Tsunewaki K (1986) The molecular basis of genetic di-
Sakamoto S (ed) Proc 6th Int Wheat Genet Symp. Maruzen,
versity among cytoplasms of Triticum and Aegilops. 5. Mito-
chondrial genome diversity among Aegilops species having
Kimber G, Tsunewaki K (1988) Genome symbols and plasma
identical chloroplast genomes. Theor Appl Genet 73:175—181
types in the wheat group. In: Miller TE, Koebner RMD (eds)
Terachi T, Tsunewaki K (1992) The molecular basis of genetic di-
Proc 7th Int Wheat Genet Symp. Bath Press, Bath, UK, pp
versity among cytoplasms of Triticum and Aegilops. VIII. Mi-
tochondrial RFLP analyses using cloned genes as probes. Mol
Lilienfeld FA (1951) H. Kihara: Genome-analysis in Triticum and
Aegilops. X. Concluding review. Cytologia 16:101–123
Tsunewaki K (1996) Plasmon analysis as the counterpart of ge-
Liu YG, Mori N, Tsunewaki K (1990) Restriction fragment length
nome analysis. In: Jauhar PP (ed) Methods of genome analysis
polymorphism (RFLP) analysis in wheat. I. Genomic DNA li-
in plants. CRC Press, New York, pp 271–299
brary construction and RFLP analysis in common wheat. Jpn J
Tsunewaki K, Wang GZ, Matsuoka Y (1996) Plasmon analysis of
Triticum (wheat) and Aegilops. 1. Production of alloplasmic
Lonsdale DM, Hodge TP, Fauron CMR (1984) The physical map
common wheats and their fertilitis. Genes Genet Syst 5:293–311
and organization of the mitochondrial genome from the fertile
Vedel F, Quetier F, Dosba F, Doussinault G (1978) Study of wheat
cytoplasm of maize. Nucleic Acids Res 12:9249–9261
phylogeny by EcoRI analysis of chloroplastic and mitochon-
Mori N, Liu YG, Tsunewaki K (1995) Wheat phylogeny deter-
mined by RFLP analysis of nuclear DNA. 2. Wild tetraploid
Vedel F, Quetier F, Cauderon Y, Dosba F, Doussinault G (1981)
Studies on maternal inheritance in polyploid wheats with cyto-
Nugent JM, Palmer JD (1991) RNA-mediated transfer of the gene
plasmic DNAs as genetic markers. Theor Appl Genet
coxII from the mitochondrion to the nucleus during flowering
Wang GZ, Miyashita NT, Tsunewaki K (1997) Plasmon analyses
Ogihara Y, Tsunewaki K (1988) Diversity and evolution of chloro-
of Triticum (wheat) and Aegilops: PCR-single-strand confor-
plast DNA in Triticum and Aegilops as revealed by restriction
mational polymorphism (PCR-SSCP) analyses of organellar
fragment analysis. Theor Appl Genet 76:321–332
DNAs. Proc Natl Acad Sci USA 94:14570–14577
Ohsako T, Wang GZ, Miyashita NT (1996) Polymerase chain re-
Watanabe N, Nakazono M, Kanno A, Tsutsumi N, Hirai A (1994)
action-single strand conformational polymorphism analysis of
Evolutionary variations in DNA sequences transferred from
intra- and interspecific variations in organellar DNA regions
chloroplast genomes to mitochondrial genomes in the Gramin-
of Aegilops mutica and related species. Genes Genet Syst
Zhang HB, Dvorak J (1992) The genome origin and evolution of
Palmer JD, Herbon LA (1986) Tricircular mitochondrial genomes
hexaploid Triticum crassum and Triticum syriacum determined
of Brassica and Raphanus: reversal of repeat configurations
from variation in repeated nucleotide sequences. Genome
by inversion. Nucleic Acids Res 14:9755–9764
Kerman university of medical sciences, kerman, iran. http://www.kmu.ac.irAntimicrobial resistance pattern of Escherichia coli causing urinary tractinfections, and that of human fecal flora, in the southeast of Iran Author(s): Mansouri, S (Mansouri, S); Shareifi, S (Shareifi, S) Source: MICROBIAL DRUG RESISTANCE-MECHANISMS EPIDEMIOLOGY AND DISEASE Volume: ٨ Issu
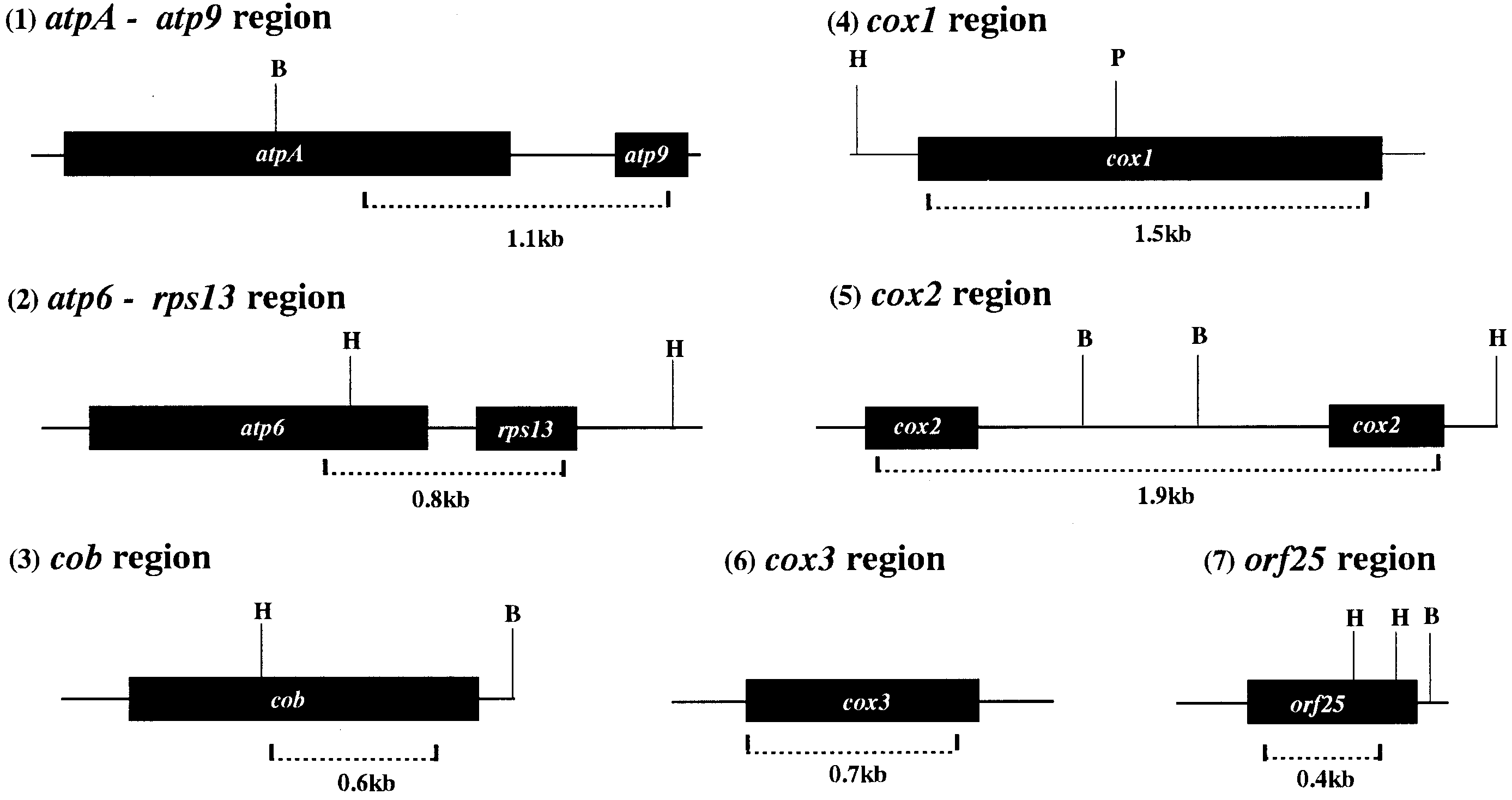 Fig. 1 Physical maps of the seven mitochondrial gene regions
Fig. 1 Physical maps of the seven mitochondrial gene regions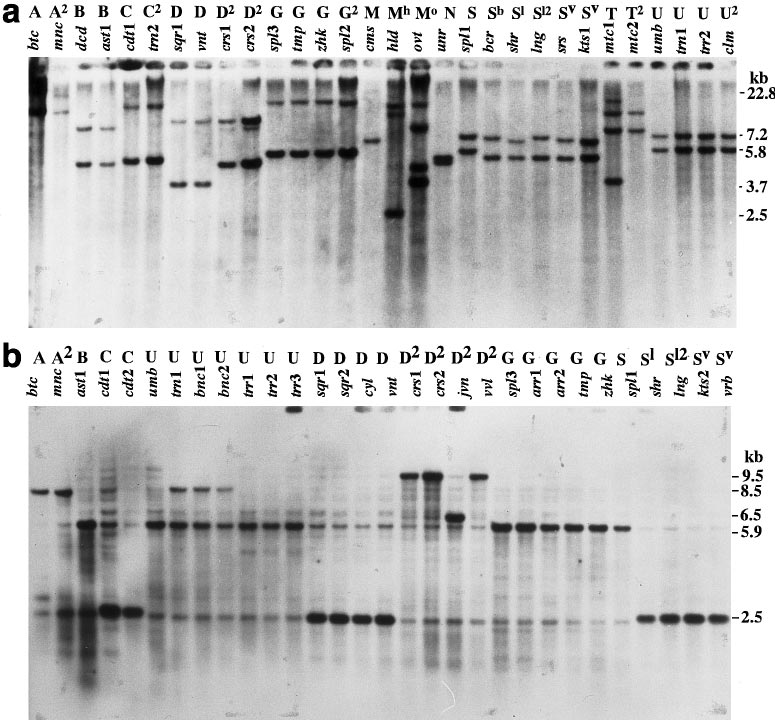 Fig. 2 a, b Southern hybridiza-
Fig. 2 a, b Southern hybridiza-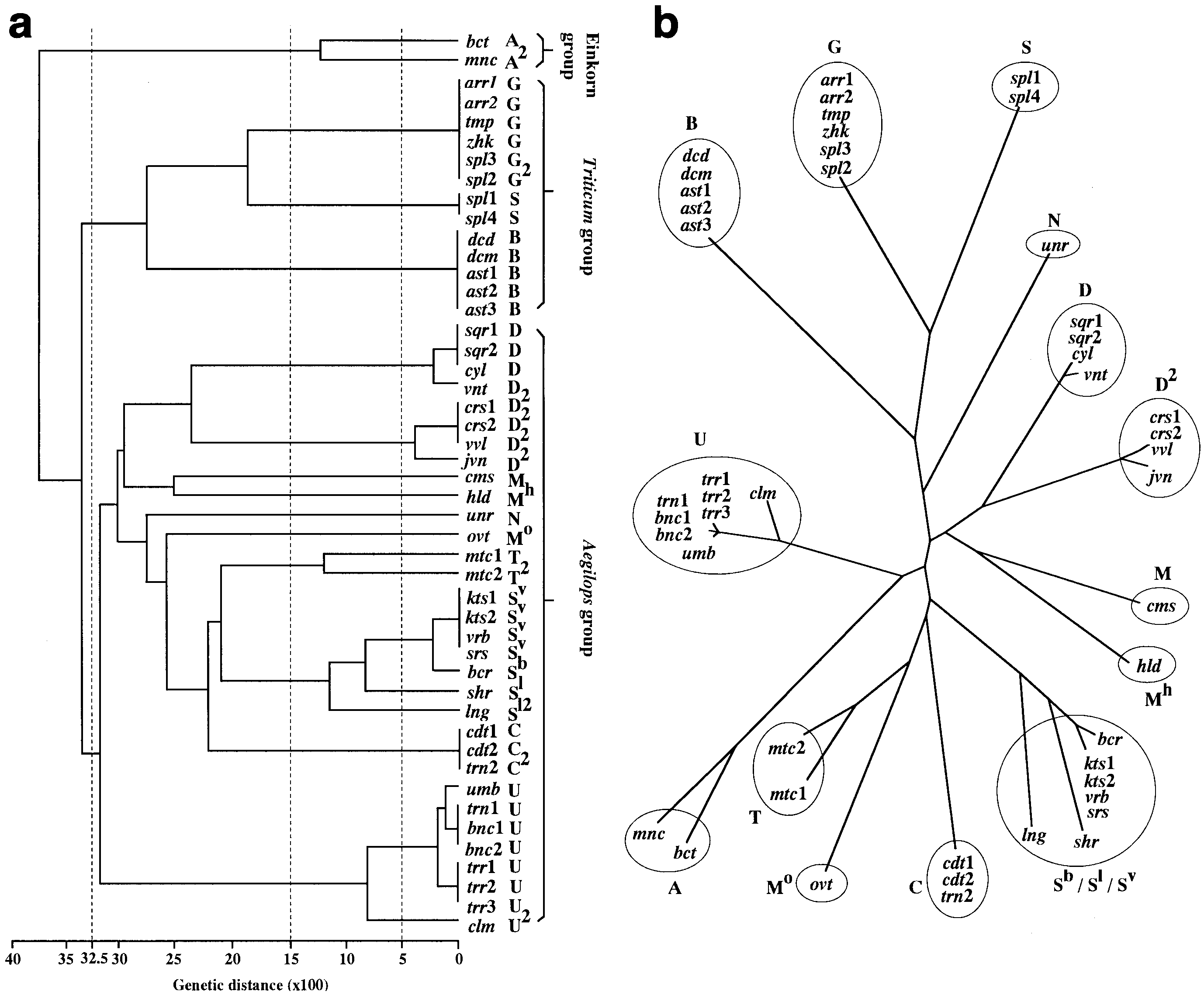 Fig. 3 a, b UPGMA (a) and NJ (b) trees for 47 chondriomes of
Fig. 3 a, b UPGMA (a) and NJ (b) trees for 47 chondriomes of