Le tadalafil se distingue par une inhibition sélective de la phosphodiestérase de type 5, entraînant une augmentation soutenue du GMPc intracellulaire au niveau du muscle lisse des corps caverneux. Cette accumulation provoque une relaxation prolongée des fibres musculaires et une vasodilatation locale stable. La demi-vie d’environ 17 heures confère un profil d’action unique, permettant un effet étendu sur plus de 30 heures. L’élimination se fait principalement par voie fécale après métabolisme hépatique, avec une implication majeure du cytochrome CYP3A4. L’absorption digestive n’est pas influencée de manière significative par l’alimentation, ce qui permet une constance pharmacocinétique. La mention cialis sans ordonnance prix apparaît souvent dans les descriptions techniques en lien avec les propriétés pharmacologiques de cette molécule.
Anneclairepannier.free.fr
Role of speed vs. grade in relation to muscle pumpfunction at locomotion onset
DON D. SHERIFF AND AMY L. HAKEMANDepartment of Exercise Science, University of Iowa, Iowa City, Iowa 52242
Received 20 July 2000; accepted in final form 1 March 2001
Sheriff, Don D., and Amy L. Hakeman. Role of speed
transduction of vasodilator chemicals (4, 16). The rel-
vs. grade in relation to muscle pump function at locomotion
ative contribution of each of these two mechanisms is
onset. J Appl Physiol 91: 269–276, 2001.—We sought to
unclear. On the basis of the known delay in the onset of
clarify the roles of contraction frequency (speed) and contrac-
arteriolar vasodilation in response to electrically in-
tion force (grade) in the rise in muscle blood flow at the onset
duced twitch contractions in anesthetized animals (6,
of locomotion. Shoemaker et al. (Can J Physiol Pharmacol 76:
12), Sheriff and co-workers (18) reasoned that the on-
418–427, 1998) explored this relationship in human hand-
set of locomotion might provide a short window of
grip exercise and found that the time course of the rise inmuscle vascular conductance was similar when a light
opportunity before vasodilation occurs in which to eval-
weight was lifted in a fast cadence and a heavy weight was
uate the isolated influence of the muscle pump on
lifted in a slow cadence (total work constant). This indicates
muscle blood flow. This possibility is supported by the
that muscle pumping (contraction frequency) was of limited
observation that the isolated muscle pump can elicit
importance in governing the time course. Rather, vasodilator
immediate increases in muscle blood flow in a setting
substances released in proportion to the total work per-
where vasodilation alone would be ineffective in rais-
formed appeared to determine the pattern and extent of the
ing flow (19). On the basis of the biphasic pattern of
rise in conductance. We hypothesized that conductance
rise in muscle vascular conductance at the onset of
would rise faster during locomotion at a high speed (fre-
mild treadmill locomotion in dogs with autonomic
quency) and low grade (force) than at a low speed and high
blockade, Sheriff and co-workers (18) attributed the
grade, despite similar total increases in conductance, owing
immediate rise in calculated vascular conductance in
to more effective muscle pumping at faster contraction rates.
response to mild exercise to the action of the muscle
Seven male rats performed nine 1-min bouts of treadmill
pump. A second, delayed rise in conductance was at-
locomotion across a combination of three speeds (5, 10, and20 m/min) and three grades (Ϫ10, 0, and ϩ15°) in random
tributed to the action of vasodilator substances, lead-
order. Locomotion at 10 m/min and 0° grade and 20 m/min
ing to an increase in vessel diameter. Consistent with
and Ϫ10° grade led to an equal rise in terminal aortic vas-
this idea, they observed in a limited number of animals
cular conductance. However, the equal rise was achieved
that a doubling of treadmill speed led to an approxi-
more quickly at the higher running speed, suggestive of more
mate doubling of the rise in vascular conductance
effective muscle pumping. Across the nine combinations of
achieved in the first 2–3 s of locomotion. That is, a
exercise, speed began to exert a statistically significant in-
doubling of contraction frequency appeared to lead to a
fluence on conductance by the 3rd s of locomotion. Grade did
proportional increase in the effectiveness of muscle
not begin to exert an influence until the 12th s of locomotion
pumping. In response to moderate exercise, conduc-
(similar to the delays reported for arteriolar dilation to mus-
tance rose smoothly (monotonically) to steady-state
cle contraction). Additional experiments in dogs provided
values, suggesting that vasodilation is initiated after a
similar results. Thus the muscle pump appears to initiate the
far shorter delay when work rate is increased above a
increase in blood flow in proportion to contraction frequency
mild intensity (no window of opportunity for evaluat-
dog; rat; nitric oxide; muscle blood flow; iliac artery; terminal
Recently, Shoemaker et al. (21) explored the rela-
aorta; arterial pressure; vasodilation; vascular conductance
tionship between muscle pumping and vasodilation atthe onset of handgrip exercise performed by humansubjects. In separate trials, these investigators had
THE ONSET OF LOCOMOTION and most forms of dynamic
subjects lift and lower a light weight in a fast cadence
exercise are accompanied by a rapid increase in the
and a heavy weight in a slow cadence, such that the
blood flow to the muscles engaged in producing move-
total work performed was equal in the two conditions.
ment. The rise in blood flow is largely attributable to a
They found that the time course of the rise in vascular
rise in the calculated vascular conductance across mus-
conductance was similar between the two conditions,
cle, which in turn is attributable to the muscle pump
indicating that muscle pumping was of limited impor-
(10, 15, 18, 23) and inhibition of arteriolar smoothmuscle after the production, release, diffusion, and
The costs of publication of this article were defrayed in part by the
payment of page charges. The article must therefore be hereby
Address for reprint requests and other correspondence: D. D.
marked ‘‘advertisement’’ in accordance with 18 U.S.C. Section 1734
Sheriff, 518 Field House, UI Exercise Science, Iowa City, IA 52242.
8750-7587/01 $5.00 Copyright 2001 the American Physiological Society
tance in governing the time course of the rise in con-
serted centrally into the vessel under isoflurane anesthesia
ductance. Rather, vasodilator substances released in
for measurement of systemic arterial pressure. Although this
proportion to the total work performed appeared to be
procedure likely reduced the blood flow to the distal tissues,
the dominant mechanism responsible for the pattern
even for the brief duration of low-intensity exercise employed
and extent of the increase in muscle vascular conduc-
in the present study, it is unlikely that the pattern (timecourse) of blood flow response measured at the level of the
terminal aorta would be substantially altered by this proce-
In the present study, we sought to pursue the follow-
dure. The catheter was tunneled to an exit site on the back.
ing two goals. First, we sought to clarify the relative
The animal was allowed to recover for Ն3 h or overnight.
role of speed (contraction frequency) vs. grade (contrac-
Data collection. The animal was lightly anesthetized (1%
tion force) in governing the rise in muscle blood flow at
isoflurane), the catheter was connected to a pressure trans-
the onset of locomotion. The rationale was that differ-
ducer (model P10EZ, Ohmeda, Madison, WI), and the flow
ences in contraction frequency would lead primarily to
transducer was connected to a flowmeter (model T106, Tran-
changes in the effectiveness of the muscle pump (at
sonic). The animal was then placed in the treadmill and, after
least in the first few seconds of locomotion), whereas
regaining full consciousness, was allowed Ն30 min to re-
differences in contraction force would lead primarily to
cover. The pressure transducer was connected to a signalconditioner (model 6600, Gould Instrument Systems, Valley
changes in arteriolar diameter [perhaps only after
View, OH). Signals were displayed on a chart recorder (model
some delay (25)]. On the basis of these premises, alter-
MT95K2, Astro-Med, West Warwick, RI), digitized at 1 kHz,
ations in speed and grade should provide insight into
and written to a fixed disk of a microcomputer with the use of
the relative roles of the muscle pump vs. vasodilation
commercially available software (PONEMAH Physiology
in raising muscle blood flow at locomotion onset. Be-
Platform, P3, Gould Instrument Systems).
cause the blood flow-raising action of the muscle pump
Experimental protocols. The animals performed nine 1-min
is proposed to be most effective during locomotory
bouts of treadmill exercise across a combination of three
exercise (10), we deemed that it would be useful to
speeds (5, 10, and 20 m/min) and three grades (Ϫ10, 0, and
apply the approach employed by Shoemaker et al. (21)
ϩ15°) in random order. For the high grade, two rats ran at
in their handgrip study to locomotory exercise. We
ϩ10° and five rats ran at ϩ15°; the group mean data arereported as ϩ15° for convenience. The stride frequency for
hypothesized that conductance would rise faster dur-
rats at these three speeds is ϳ1, 2, and 3 strides/s (3). The
ing locomotion at a high speed and low grade than at a
animals were allowed to recover for Ն3 min between bouts.
low speed and high grade, despite similar total in-
Data analysis. Terminal aortic vascular conductance was
creases in conductance, owing to more effective muscle
calculated as terminal aortic flow divided by arterial pres-
pumping at faster contraction rates associated with
sure. Arterial pressure, terminal aortic flow, and terminal
higher treadmill speeds. Second, because nitric oxide
aortic vascular conductance were each averaged over 1-s
(NO) synthase (NOS) inhibition has recently been re-
periods beginning 10 s before the onset of locomotion until
ported to slow the vasodilation in response to locomo-
tion (17), we inhibited NOS in an effort to extend theshort window of opportunity for evaluating muscle
pump function at the onset of locomotion at moderate
Six mongrel dogs (18–24 kg body wt) of either gender were
intensities of locomotion. Experiments were carried
selected for their willingness to run on a motor-driven tread-
out in chronically prepared, conscious rats and dogs.
mill (model J6, Proform). The dogs were familiarized withtreadmill running in a series of training sessions before the
following aseptic surgical procedures were performed. Surgical preparation. Dogs were anesthetized with thio-
All procedures met National Institutes of Health guide-
pental, intubated, ventilated, and maintained with halo-
lines and were reviewed and approved by the InstitutionalAnimal Care and Use Committee of the University of Iowa.
thane. Ultrasonic transit-time blood flow transducers (Tran-sonic) and vascular occluder cuffs were placed bilaterally on
the iliac arteries through a midline abdominal incision. Inone dog, these devices were implanted on the terminal aorta,
Seven male Sprague-Dawley rats (250–300 g) were se-
and the measured values of blood flow in this dog were on
lected for their willingness to run on a motor-driven tread-
average three times greater than in the other dogs. Blood
mill (model 1010 Modular Treadmill, Columbus Instruments,
flows measured in this dog were divided by a factor of 3 before
Columbus, OH). The rats were familiarized with treadmill
they were averaged with the results from the remaining
running before the following aseptic surgical procedures
animals. A catheter was inserted into the aorta for measure-
ment of systemic arterial pressure and in a femoral artery
Surgical preparation. Rats were anesthetized with isoflu-
and vein for measurement of hindlimb perfusion pressure
rane. An ultrasonic transit-time blood flow transducer (model
and infusion of drugs, respectively. All leads were tunneled to
1.5RB, Transonic, Ithaca, NY) was implanted in each animal
exit sites on the back. Skin patches delivering a total of 75
on the terminal aorta through a midline abdominal incision.
g/h of fentanyl were placed on the dogs for 72 h after
The probe cable was tunneled to an exit site on the back. The
surgery to control postoperative pain, and the dogs were
animal was given nalbuphine hydrochloride (1 mg/kg sc) for
given cephalexin (500 mg po twice a day) continually after
control of postoperative pain. The animal was allowed to
surgery throughout the time that data were collected.
recover until an acceptable blood flow signal was acquired
Data collection. Catheters were connected to pressure
(usually 2–3 days). On the day before or on the morning of a
transducers (model P10EZ, Ohmeda) and flow transducers
day when an experiment was to be carried out, a femoral
were connected to flowmeters (model T106, Transonic). Sig-
artery was ligated, and a PE-10 catheter was directly in-
nals were displayed on a pen recorder and digitized at 250
Hz. Average values of each signal were written to a fixed diskof a microcomputer twice per second. Experimental protocols. The animals performed treadmill
exercise at three intensities (3.2 km/h and 0% grade, 6.4km/h and 0% grade, and 6.4 km/h and 10% grade) for 3 minin no regular order. The animals were allowed to recover for
Ն3 min before exercise was repeated at a different exerciseintensity. The animals were then given intravenous hexame-thonium (10 mg/kg), atropine (0.1 mg/kg), and captopril (1mg/kg) to block autonomic function and the renin-angioten-sin system, respectively (17, 20). The efficacy of these drugswas inferred from the exaggerated fall in arterial pressurethat accompanied locomotion after autonomic blockade. Thethree bouts of exercise were then repeated. Some animals ranfor only 1 min at some workloads, owing to reduced exercisecapacity after autonomic blockade. Animals were then givennitro-L-arginine methyl ester (L-NAME, 10 mg/kg iv) to inhibitNO production, and exercise was repeated as describedabove. The efficacy of NOS inhibition was inferred from therise in arterial pressure elicited by this drug. On a separateday, animals performed locomotion as described above withand without L-NAME alone. Drugs were acquired from SigmaChemical (St. Louis, MO). Data analysis. Iliac vascular conductance was calculated
as iliac blood flow divided by arterial pressure. We evaluatedthe effect of L-NAME treatment on the time course of the risein iliac vascular conductance graphically on the basis ofcalculations of iliac vascular conductance for each exercise
Fig. 1. Hemodynamic responses from 1 rat to 9 bouts of treadmill
bout averaged over 1-s periods from 10 s before until the end
locomotion graded across 3 speeds and 3 grades. Terminal aortic flow
of the first 60 s of exercise. Values are means Ϯ SE.
and vascular conductance rose in response to locomotion, whereasfemoral arterial pressure tended to fall slightly. There was consid-erable overlap in the steady-state values of flow and conductance
Tests for statistical significance were done by paired
t-tests, except as follows. For each 1-s time period of the data
0.150 Ϯ 0.022 and 0.162 Ϯ 0.023 ml ⅐ minϪ1 ⅐ mmHgϪ1
presented in Figs. 3 and 5, treatment effects were tested
for 10 m/min and 0° grade and 20 m/min and Ϫ10°
statistically by multiple linear regression (22) with a com-
grade, respectively (P ϭ 0.19). Exercise vascular con-
puter spreadsheet program (Microsoft Excel 97, Redmond,WA). Dummy variables were used as independent variables
ductance (averaged over the final 10 s of locomotion)
to account for interindividual variability among animals (22).
was 0.271 Ϯ 0.034 and 0.277 Ϯ 0.038 ml ⅐ minϪ1 ⅐
For Fig. 3, treadmill speed (5, 10, or 20 m/min), grade (Ϫ10,
mmHgϪ1 for 10 m/min and 0° grade and 20 m/min and
0, or ϩ10°), and sequence order (1–9) were used to encode
Ϫ10° grade, respectively (P ϭ 0.60). The increase in
treatment effects. For Fig. 5, treadmill speed (3.2 or 6.4
hindlimb conductance from rest to exercise was
km/h) and treadmill grade (0 or 10%) were used to encode
0.121 Ϯ 0.016 and 0.115 Ϯ 0.021 ml ⅐ minϪ1 ⅐ mmHgϪ1
treatment effects. Values are means Ϯ SE.
for 10 m/min and 0° grade and 20 m/min and Ϫ10°grade, respectively (P ϭ 0.53). Femoral arterial pres-
sure was similar during the two bouts. Figure 2 depicts
the time course of the rise in hindlimb conductance aspercent increase, and it can be seen that the similar
Figure 1 depicts arterial pressure, hindlimb blood
increases in conductance followed different time
flow, and hindlimb vascular conductance in a rat per-
courses. That is, conductance at the slower speed and
forming treadmill exercise graded across nine combi-
higher grade was significantly less (P Ͻ 0.05) than
nations stemming from three speeds (5, 10, and 20
conductance at the faster speed and lesser grade early
m/min) and three grades (Ϫ10, 0, and ϩ15°). All speeds
and grades led to immediate increases in blood flow
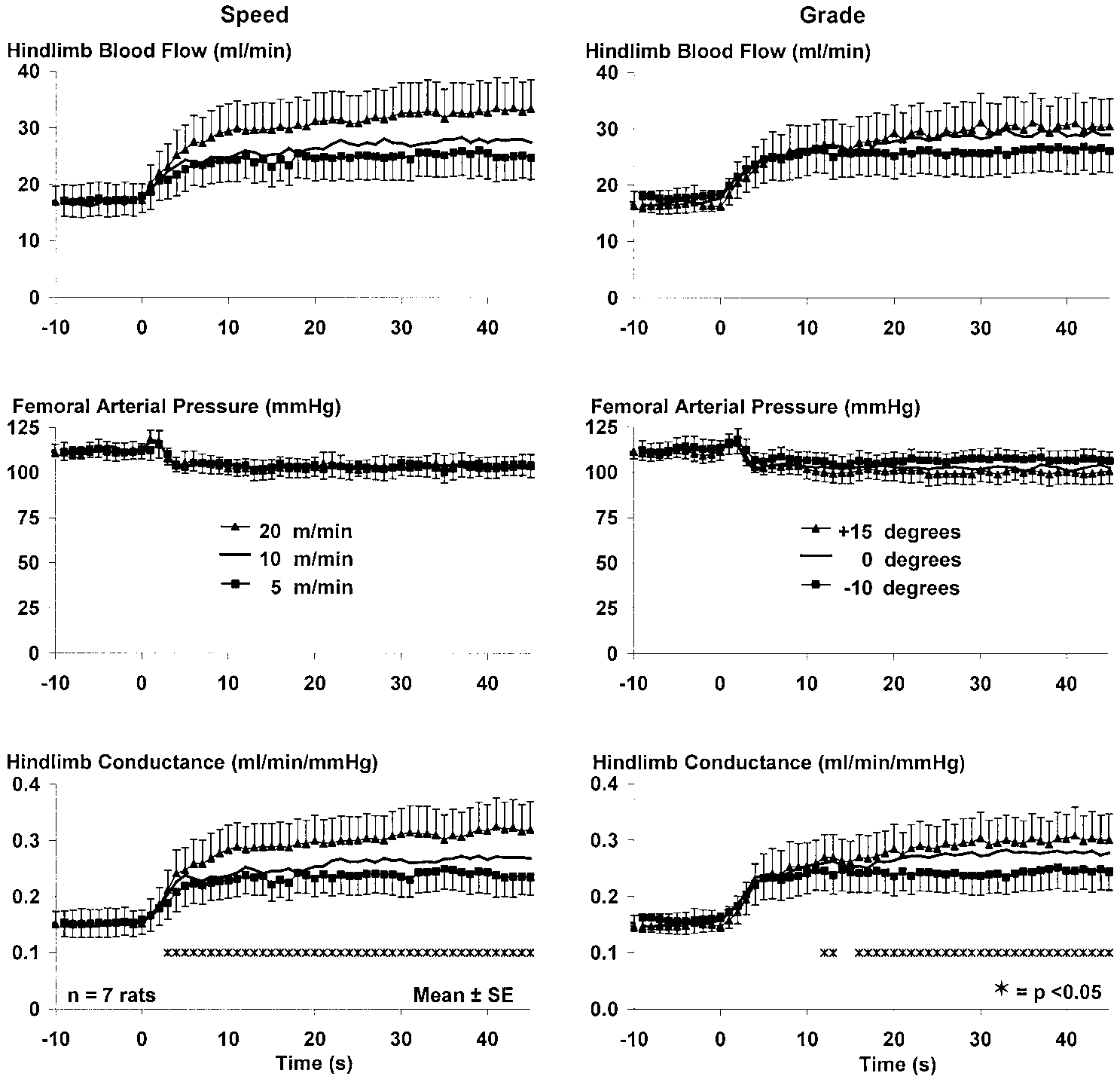
Figure 3 depicts group mean data showing the
and calculated vascular conductance. The steady-state
changes in hindlimb blood flow, femoral arterial pres-
values of these variables reveal considerable overlap
sure, and hindlimb vascular conductance from seven
among the different combinations of speed and grade.
rats. Figure 3, left, shows the isolated influence of
Data collected in response to locomotion at 10 m/min
speed on the rise in blood flow and conductance; i.e.,
and 0° grade and 20 m/min and Ϫ10° grade were
each trace represents the response to the three indi-
selected to illustrate how different combinations of
vidual bouts at the three different grades, averaged
speed and grade can lead to a similar rise in vascular
together. The statistical analysis (which included the
conductance with a variable pattern of response. Rest-
data from the intermediate speed) indicates that speed
ing vascular conductance (averaged over the 10 s im-
began to exert a significant influence (P Ͻ 0.05) on
mediately preceding the onset of locomotion) was
conductance as soon as the 3rd s of locomotion, and this
together. The statistical analysis (which included thedata from the intermediate grade) indicates that gradedid not begin to exert a significant effect (P Ͻ 0.05) onthe rise in conductance until the 12th s of locomotion.
Figure 4 depicts the time course of the rise in hind-
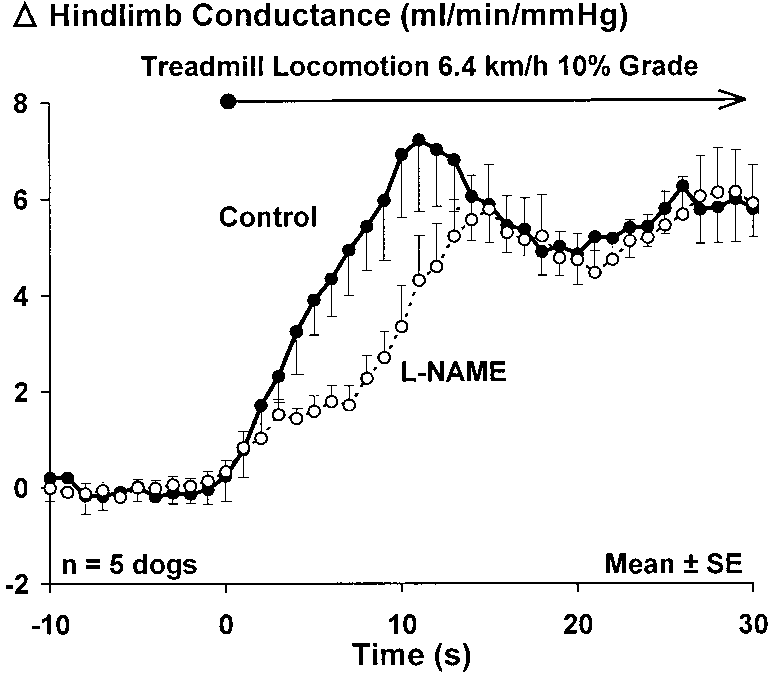
limb vascular conductance averaged from five dogsperforming treadmill exercise at 6.4 km/h and 10%grade before and after inhibition of NOS (autonomicfunction intact). When NOS function was intact, con-ductance rose progressively over the first 10 s of loco-motion with an apparent overshoot. After inhibition ofNOS, the time course of the rise in conductance exhib-ited a biphasic pattern. Conductance initially (1–3 s)
Fig. 2. Time course of rise in hindlimb vascular conductance as
rose to a steady level that persisted for ϳ5 s, after
percent increase in response to locomotion at 10 m/min and 0° grade
which conductance began a second rise to a steady-
and 20 m/min and Ϫ10° grade in rats. Resting vascular conductance,
state value with no apparent overshoot.
exercise vascular conductance, and change in vascular conductance
Figure 5 depicts the rise in conductance across the
from rest to exercise were not significantly different between the 2conditions. Rise in vascular conductance follows a slower time course
three workloads after inhibition of NOS and autonomic
during locomotion at 10 m/min and 0° grade than during locomotion
blockade. The immediate rise in conductance in the
first few seconds of locomotion at 6.4 km/h was approx-imately twice as great as the rise that accompanied
influence persisted throughout the remainder of exer-
locomotion at 3.2 km/h. Furthermore, the immediate
cise. Figure 3, right, depicts the isolated influence of
rise in conductance was identical for locomotion at 6.4
grade on the rise in blood flow and conductance; i.e.,
km/h and 0% grade and for locomotion at 6.4 km/h and
each trace represents the response to the three indi-
10% grade. Thereafter, under all three conditions, con-
vidual bouts at the three different speeds, averaged
ductance underwent further increases.
Fig. 3. Isolated influence of speed andgrade on the hemodynamic response tolocomotion in rats. Left: each trace rep-resents responses to individual bouts atthe 3 different grades, averaged to-gether. Right: each trace represents re-sponses to individual bouts at the 3 dif-ferent speeds at a single grade, averagedtogether. Error bars for intermediatespeed and grade have been deleted toimprove clarity.
characteristics, which are relatively slow (half-relax-ation times of 5–40 s) (11). Therefore, we reasoned thata fixed rise in vascular conductance should follow asimilar time course, regardless of the specific mechan-ics of locomotion if vasodilation were the sole or over-whelming cause of the rise in conductance. In supportof this rationale is the observation that vasodilatoryresponses induced by four different chemical sub-stances follow a relatively similar time course (25). Also, even though a change in arteriolar diameter canbe detected immediately after an electrically induced1-s tetanic contraction of surrounding skeletal musclefibers, the vasodilation that is expressed lasts for ϳ50s (6). These observations imply that the microvascula-ture possesses relatively sluggish response character-
Fig. 4. Influence of nitric oxide synthase (NOS) inhibition on the
istics and thus functions as a low-pass filter.
time course of the rise in hindlimb vascular conductance in response
The muscle pump exerts important influences on the
to locomotion of moderate intensity in dogs. NOS inhibition elimi-
pressure-volume (capacitive) (20) and the pressure-
nated the rise in conductance normally seen at 3–7 s but did not
flow characteristics (18, 19) of the peripheral circula-
affect the overall rise in conductance. Nitro-L-arginine methyl ester(L-NAME) exerted a statistically significant effect (P
tion. A number of investigators employing a broad
s. Resting conductance was 2.60 Ϯ 0.19 and 1.23 Ϯ 1.23
mixture of different exercise conditions have concluded
ml ⅐ minϪ1 ⅐ mmHgϪ1 in control and after L-NAME, respectively.
that the muscle pump can augment blood flow acrossmuscle (5, 15, 18, 19, 23). Laughlin (10) postulated that
DISCUSSION
the blood flow-raising function of the muscle pump ismost effective during locomotory-type exercise. A com-
The major new findings of our study are twofold.
mon assumption is that contraction frequency consti-
First, the time course of the rise in muscle vascular
tutes a major determinant of muscle pump efficacy (5,
conductance is significantly altered by the mechanical
8, 18, 21) just as cardiac frequency can constitute a
factors (speed and grade) by which a given rate of work
major determinant of cardiac pump efficacy, and sev-
is achieved during locomotion in rats. Second, aug-
eral studies have provided evidence in support of this
mented NO formation exerts a significant effect on the
idea (5, 8, 18). For example, Gotschall and co-workers
rise in conductance during the initial stages (3–10 s) in
(8) examined the importance of cycling cadence at a
response to a moderate intensity of treadmill locomo-
fixed workload and found that higher cadences were
tion and appears to account for all the dilation that
associated with a higher total vascular conductance,
normally occurs over this period in dogs. Moreover, the
which they attributed to more effective muscle pump-
absence of the normal influence of NO reveals that themuscle pump likely accounts for all the rise in bloodflow in the first 8 s of locomotion in these conditions.
The increases in muscle blood flow and muscle vas-
cular conductance achieved during dynamic exerciseare tightly coupled to the amount of work performed bymuscle, and metabolic vasodilation is an importantregulatory mechanism by which muscle blood flow iscoupled to local energy demands. The common view isthat vasodilator substances within active muscle accu-mulate in a manner governed by the balance betweenthe energy expended by the muscle and the blood flowthrough the muscle and that metabolic vasodilationconstitutes the primary determinant of the vascularconductance achieved during locomotion. Because ar-terial pressure changes far less than does muscle bloodflow in the transition from rest to locomotion (20% vs.
Fig. 5. Rise in hindlimb vascular conductance in response to loco-
200%), the blood flow achieved is also tightly coupled
motion graded across 2 speeds and 2 grades after inhibition of NOS
to the rise in vascular conductance. For these reasons,
and autonomic blockade. Doubling of treadmill speed from 3.2 to 6.4
we chose muscle vascular conductance as an index of
km/h led to an approximate doubling of the rise in conductance
the work rate imposed by the various combinations of
achieved in the first 5 s of locomotion. Increasing treadmill grade
speed and grade employed in our study. Also, the time
from 0 to 10% at a fixed treadmill speed did not affect the rise invascular conductance over this time period. Speed began exerting a
course of the relaxation of vascular smooth muscle
significant effect (P Ͻ 0.05) on conductance at 4 s, whereas grade did
appears to be constrained by relatively fixed dynamic
not begin to exert an effect until 18 s.
ing. The importance of contraction force on the bloodflow-raising function of the muscle pump is unclear,although this variable is known to be relatively unim-portant in determining the efficacy of the muscle pumpin altering the pressure-volume characteristics of thecirculation [i.e., mild contractions are as effective asforceful contractions in emptying muscle veins (2)].
We found that contraction frequency exerts an influ-
ence on muscle vascular conductance as soon as the 3rds of locomotion. In contrast, the influence of musclecontraction frequency on arteriolar dilation per se isindistinct. For example, the changes in arteriolar di-ameter in response to contractions at 2 and 4/s (similarto the contraction frequencies of the rats in the presentstudy) are identical for the first 40 s, whereas the risein diameter is more extensive over this period during
Fig. 6. Influence of speed and grade on the rise in hindlimb vascular
contractions at 8/s (6). On the basis of this finding, the
conductance in response to locomotion in rats. Results are averaged
observation that conductance is different across vari-
together over time periods when a factor (speed or grade) does notexert a statistically significant influence, and the results are split
ous treadmill speeds relatively early during locomotion
over time periods when a factor does exert a significant influence.
argues that this difference stems from the influence ofcontraction frequency on the effectiveness of musclepumping, and not on arteriolar vasodilation.
gether into a single trace. At the point in time when
We found that treadmill grade (contraction force) did
speed begins to exert a significant influence (3rd s), the
not begin to exert a significant effect on conductance
single trace splits into three traces, one new trace for
until 13 s after the onset of locomotion. If contraction
each of the three speeds. Finally, at the point in time
force constituted an important determinant of the effi-
when grade begins to exert a significant influence (12th
cacy of muscle pumping, we would expect this variable
s), the traces representing locomotion at the low and
to exert an effect on conductance with little or no time
high speeds are each split into three, one new trace for
delay. The delayed nature of the influence of contrac-
each of the three grades. The responses from the three
tion force on conductance is consistent with the idea
grades at the intermediate speed remain averaged
that this factor exerts its influence by modulating the
together to improve clarity. As can be seen, a similar
release of vasodilator substances. The fact that the
rise in conductance can be achieved via many paths,
delay we report here is similar to the delays reported
for arterioles to begin dilating in response to twitch
The qualitatively different results from the study by
contractions at similar frequencies strengthens this
Shoemaker et al. (21) and the present study likely stem
from the different modes of exercise employed in the
Recently, Shoemaker et al. (21) found that contrac-
two studies. In the study by Shoemaker et al., the
tion frequency did not affect the pattern or extent of
subjects raised and then lowered a weight over a 1-s
the rise in muscle blood flow during forearm exercise
period and then relaxed the muscle for 1 s before
performed by human subjects. Rather, the rise in mus-
another duty cycle was begun. Thus for each duty cycle,
cle blood flow during work at a fixed rate followed the
the muscle was continuously activated for 1 s, during
same time course, regardless of whether the work was
which it performed a concentric contraction followed by
performed with a light weight at a fast cadence or with
an eccentric contraction, after which it relaxed at fixed
a heavy weight at a slow cadence. This led these inves-
length. In contrast, the pattern of activation and the
tigators to conclude that vasodilator responses are im-
mode of shortening and lengthening are substantially
portant early during dynamic exercise, and this influ-
different during locomotion. Generally, muscles en-
ence likely obscures any muscle pump effect that is
gaged in producing locomotion tend to perform brief
dependent on contraction frequency. In contrast, we
concentric contractions, relax, and then undergo forced
found that the time course of the rise in vascular
reextension by antagonistic muscles while they are
conductance varied significantly with changes in con-
relaxed (or are relaxing). For example, during locomo-
traction rate and grade, even when the eventual rise in
tion at 8.5 km/h (2.2 strides/s) in dogs, the gluteus
vascular conductance, and thus presumably the total
medius muscle (a hip extensor) expresses electromyo-
work rate, was the same (Fig. 2). Furthermore, the
genic activity for 27% (120 ms) of the stride period and
faster rise in conductance at the faster treadmill speed
the muscle shortens while it is active (7). The muscle is
is consistent with the idea that the muscle pump is
then quiescent for Ͼ150 ms before it begins to re-
more effective at higher contraction rates. To illustrate
lengthen as the leg is pulled forward by the hip flexors.
these points graphically, Fig. 6 replots the vascular
For this muscle to accumulate 1 s of electromyographic
conductance data from the nine bouts of exercise as
activity (as presumably occurs in the 1st duty cycle in
follows. Over the period of time when neither speed nor
the study of Shoemaker et al.), it would take over eight
grade exerted a significant influence (from the begin-
strides and thus would not occur until the 4th s of
ning until the 3rd s), all the runs were averaged to-
exercise. The forceful reextension of muscle may be a
critical factor in promoting effective muscle pumping
in time, an observation that underscores the view that
during locomotion (10). Histological evaluation of the
redundant control mechanisms are involved in the
influence of changes in muscle length on lymphatic
regulation of muscle blood flow (9).
volume (13) and microscopy evaluation of the influence
In the present study, we confirm and extend the
of changes in muscle length on the geometry of mi-
previous findings of Sheriff et al. (18). Figure 5 shows,
crovessels (14) lend credence to the suggestion that
on the basis of the measurement of iliac artery blood
skeletal muscle and its vasculature function as a “bel-
flow and without the competing influence of the auto-
nomic nervous system and the early contribution ofNO, that an approximate doubling of stride frequency
(treadmill speed) leads to an approximate doubling ofthe initial (2- to 3-s) rise in conductance. Importantly,
In a previous study using a limited number of ani-
when the influence of NO is lacking, increasing tread-
mals, Sheriff et al. (18) found that doubling treadmill
mill grade at a fixed speed does not alter the initial (1-
speed from a low to a moderate level led to a doubling
to 10-s) rise in conductance. Thus all the other numer-
of the immediate rise in hindlimb conductance in dogs
ous potential vasodilator substances (16) fail to elicit
walking on the flat. These investigators attributed this
any measurable change in conductance until ϳ8 s into
initial (2- to 3-s) rise in blood flow and calculated
the exercise bout. Beyond 8 s, these other dilators
conductance to more effective muscle pumping at the
eventually exert a profound effect, eliciting a second
higher contraction frequency. After a delay of ϳ10 s,
rise in conductance that is approximately twice as
during which time conductance was relatively un-
large for work at 10% grade as on the flat. In our
changed, they observed that conductance underwent a
opinion, it is unlikely that vasodilation causes (or even
second rise that they attributed to the action of vaso-
contributes to) the initial (1- to 2-s) rise in conductance
dilator substances. They also found that vascular con-
under these conditions, and it is even more unlikely
ductance rose more smoothly to its steady-state level in
that vasodilation ceases for 5 s and then begins
dogs walking uphill at a moderate speed, suggesting
again. For example, recent elegant studies by Welsh
that vasodilation might begin much sooner at this
and Segal (24) provide compelling evidence that
workload (thereby encroaching on the short window of
there is a 2- to 3-s delay attributable to electrome-
opportunity for gauging the effectiveness of the muscle
chanical coupling within vascular smooth muscle
pump). Importantly, recent evidence indicates thatNOS inhibition slows the rate of vasodilation during
cells in response to vasodilator substances. Also,
locomotion (17). It is believed that the muscle pump
Wunsch et al. (25) recently demonstrated a delay of
elicits an immediate increase in blood flow and, as a
ϳ4 s before the onset of dilation of isolated arterioles
consequence, there is a rise in shear stress on endo-
after direct application of chemical vasodilator sub-
thelial cells, which augments the release of NO. In
stances. On the basis of these arguments, our results
this way, NO acts as an amplifier that reinforces the
support the proposal of Sheriff et al. that the muscle
influence of the muscle pump on blood flow. In the
pump elicits all the initial (1- to 3-s) increase in
present study, we sought to shut off this amplifier by
calculated conductance (blood flow) during mild in-
inhibiting NOS as a means of possibly extending the
tensities of locomotion and extends this to moderate
window of opportunity for evaluating muscle pump
intensities of locomotion when NO function is inhib-
effectiveness at the onset of locomotion at this higher
ited. The discrepancy between the delayed vasodila-
tory responses resulting from direct application of
We found that NO exerts an important influence
vasodilator substances to isolated arterioles (25) and
early during the transition from quiet standing to
the more immediate vasodilation observed in arte-
locomotion as reported previously (17). The focus of our
rioles after 1-s electrically evoked contractions of
previous study was on evaluating the contribution of
surrounding skeletal muscle (6) remains to be ex-
NO to the rise in vascular conductance that accompa-
nies locomotion; the focus here is on evaluating what
Finally, our results may help explain why the search
the absence of NO reveals about muscle pump function.
for the chemical substances that account for active
As seen in Fig. 4, NOS inhibition does not alter the
hyperemia has proven so elusive (9). Our results
initial (1- to 2-s) rise in conductance. However, NOS
strongly suggest that the muscle pump can initiate a
inhibition appears to eliminate the entire rise in con-
significant (2-fold) increase in blood flow, which in turn
ductance normally seen in seconds 3–7 at this intensity
stimulates augmented NO release and a further rise in
of locomotion. These results are consistent with the
blood flow. If feedforward control such as this matches
idea that the muscle pump initiates an immediate
the rise in blood flow to the rise in oxygen demand in a
increase in blood flow (virtual conductance) that is
close temporal relationship, a chemical error signal
subsequently reinforced by augmented NO formation
may not arise over a wide range of intensities of loco-
induced by an increase in endothelial wall shear stress
motion. Indeed, the tendency for vascular conductance
that arises as a consequence of the increase in blood
to exhibit an overshoot early during mild locomotion
flow. Beyond 7 s, conductance begins a second rise
when sympathetic function is intact (18) indicates that
during NOS inhibition that is similar in rate and
muscle may be relatively overperfused early during
extent to the rise seen in control conditions but delayed
We thank Tony Smith for expert technical assistance.
14. Nakao M and Segal SS. Muscle length alters geometry of
This work was supported by National Heart, Lung, and Blood
arterioles and venules in hamster retractor. Am J Physiol HeartCirc Physiol 268: H336–H344, 1995.
15. Radegran G and Saltin B. Muscle blood flow at onset of
dynamic exercise in humans. Am J Physiol Heart Circ PhysiolREFERENCES
1. Almen T and Nylander G. Serial phlebography of the normal
16. Shepherd JT. Circulation to skeletal muscle. In: Handbook of
lower leg during muscular contraction and relaxation. Acta Ra-Physiology. The Cardiovascular System. Peripheral Circulationand Organ Blood Flow. Bethesda, MD: Am. Physiol. Soc., 1983,
2. Barendsen GJ and van den Berg JW. Venous capacity, ve-
sect. 2, vol. III, pt. 1, chapt. 11, p. 319–370.
17. Sheriff DD, Nelson CD, and Sundermann RK. Does auto-
nous refill time and the effectiveness of the calf muscle pump in
nomic blockade reveal a potent contribution of nitric oxide to
normal subjects. Angiology 35: 163–172, 1984.
locomotion-induced vasodilation? Am J Physiol Heart Circ
3. Clarke KA and Parker AJ. A quantitative study of normal Physiol 279: H726–H732, 2000.
locomotion in the rat. Physiol Behav 38: 345–351, 1986.
18. Sheriff DD, Rowell LB, and Scher AM. Is rapid rise in
4. Delp MD. Control of skeletal muscle perfusion at the onset of
vascular conductance at onset of dynamic exercise due to muscle
dynamic exercise. Med Sci Sports Exerc 31: 1011–1018, 1999.
pump? Am J Physiol Heart Circ Physiol 265: H1227–H1234,
5. Folkow B, Gaskell P, and Waaler BA. Blood flow through
limb muscles during heavy rhythmic exercise. Acta Physiol
19. Sheriff DD and Van Bibber R. Flow-generating capability of
the isolated skeletal muscle pump. Am J Physiol Heart Circ
6. Gorczynski RJ, Klitzman B, and Duling BR. Interrelations Physiol 274: H1502–H1508, 1998.
between contracting striated muscle and precapillary microves-
20. Sheriff DD, Zhou XP, Scher AM, and Rowell LB. Depen-
sels. Am J Physiol Heart Circ Physiol 235: H494–H504, 1978.
dence of cardiac filling pressure on cardiac output during rest
7. Goslow, GE Jr, Seeherman HJ, Taylor CR, McCutchin MN,
and dynamic exercise in dogs. Am J Physiol Heart Circ Physioland Heglund NC. Electrical activity and relative length
changes of dog limb muscles as a function of speed and gait. J
21. Shoemaker JK, Tschakovsky ME, and Hughson RL. Vaso-
dilation contributes to the rapid hyperaemia with rhythmic
8. Gotshall RW, Bauer TA, and Fahrner SL. Cycling cadence
contractions in humans. Can J Physiol Pharmacol 76: 418–427,
alters exercise hemodynamics. Int J Sports Med 17: 17–21, 1995.
9. Joyner MJ and Proctor DN. Muscle blood flow during exer-
22. Slinker BK and Glantz S. Multiple linear regression is a
cise: the limits of reductionism. Med Sci Sports Exerc 31: 1036–
useful alternative to traditional analyses of variance. Am JPhysiol Regulatory Integrative Comp Physiol 255: R353–R367,
10. Laughlin MH. Skeletal muscle blood flow capacity: role of
muscle pump in exercise hyperemia. Am J Physiol Heart Circ
23. Tschakovsky ME, Shoemaker JK, and Hughson RL. Vaso- Physiol 253: H993–H1004, 1987.
dilation and muscle pump contribution to immediate exercise
11. Liu X, Jiang H, and Stephens NL. Use of a new index to study
hyperemia. Am J Physiol Heart Circ Physiol 271: H1697–H1701,
relaxation in a vascular model of anaphylactic shock. J ApplPhysiol 74: 2621–2626, 1993.
24. Welsh DG and Segal SS. Endothelial and smooth muscle cell
12. Marshall JM and Tandon HC. Direct observations of muscle
conduction in arterioles controlling blood flow. Am J Physiol
arterioles and venules following contraction of skeletal muscle
Heart Circ Physiol 274: H178–H186, 1998.
fibres in the rat. J Physiol (Lond) 350: 447–459, 1984.
25. Wunsch SA, Muller-Delp J, and Delp MD. Time course of
13. Mazzoni MC, Skalak TC, and Schmid-Scho ¨ nbein GW. Ef-
vasodilatory responses in skeletal muscle arterioles: role in hy-
fects of skeletal muscle fiber deformation on lymphatic volumes.
peremia at the onset of exercise. Am J Physiol Heart Circ PhysiolAm J Physiol Heart Circ Physiol 259: H1860–H1868, 1990.
Attention Pastors Christians & Society TO-DAY, pages 3 and 4, can bereproduced locally on a pho-tocopier for use as a supple-ment or insert with bulletinsor newsletters. Pfizer pollutes physical, moral environments, S.C. Johnson says values may vary ANTI-CHRISTIAN Not only is Pfizer one of the tice Department covers viola-sible Television (CLeaR-TV) leading sponsors of sex, viole
THE η(1405), η(1475), f1(1420), AND f1(1510)Revised November 2013 by C. Amsler (Bern) and A. Masoni(INFN Cagliari). The first observation of the η(1440) was made in pp anni-hilation at rest into η(1440)π+π−, η(1440) → KKπ [1]. Thisstate was reported to decay through a0(980)π and K∗(892)Kwith roughly equal contributions. The η(1440) was also ob-served in radi
 Role of speed vs. grade in relation to muscle pumpfunction at locomotion onset
DON D. SHERIFF AND AMY L. HAKEMANDepartment of Exercise Science, University of Iowa, Iowa City, Iowa 52242
Received 20 July 2000; accepted in final form 1 March 2001
Sheriff, Don D., and Amy L. Hakeman. Role of speed
Role of speed vs. grade in relation to muscle pumpfunction at locomotion onset
DON D. SHERIFF AND AMY L. HAKEMANDepartment of Exercise Science, University of Iowa, Iowa City, Iowa 52242
Received 20 July 2000; accepted in final form 1 March 2001
Sheriff, Don D., and Amy L. Hakeman. Role of speed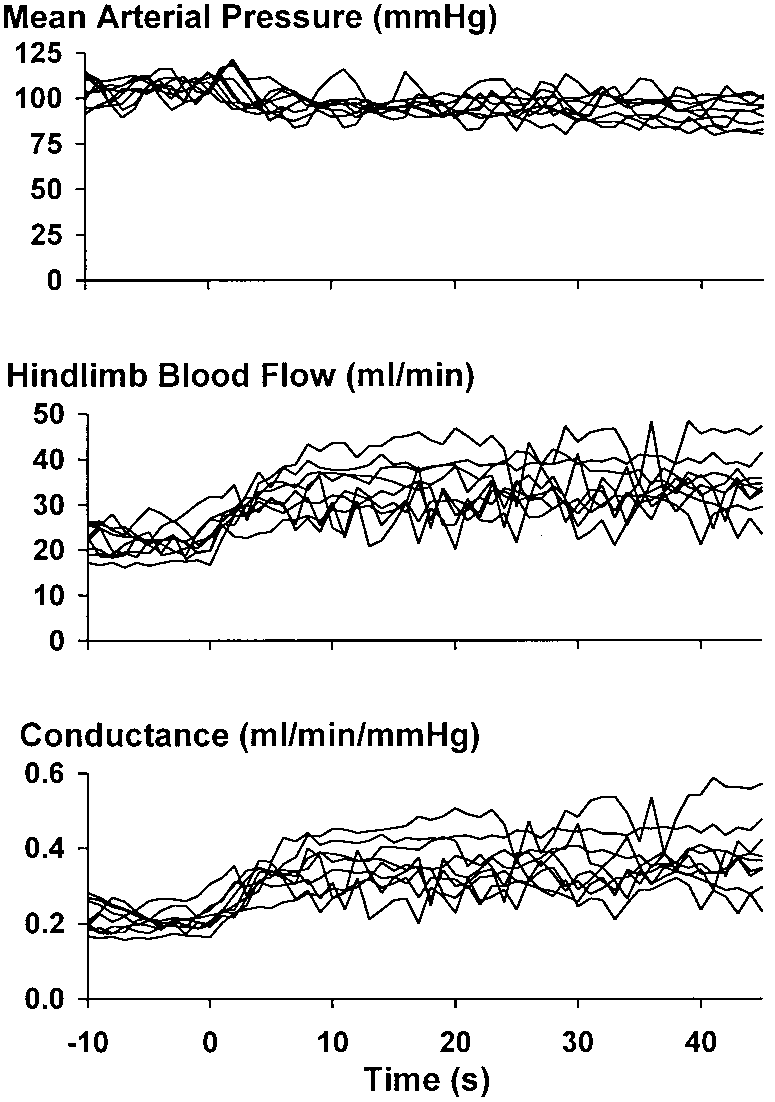 Hz. Average values of each signal were written to a fixed diskof a microcomputer twice per second.
Hz. Average values of each signal were written to a fixed diskof a microcomputer twice per second.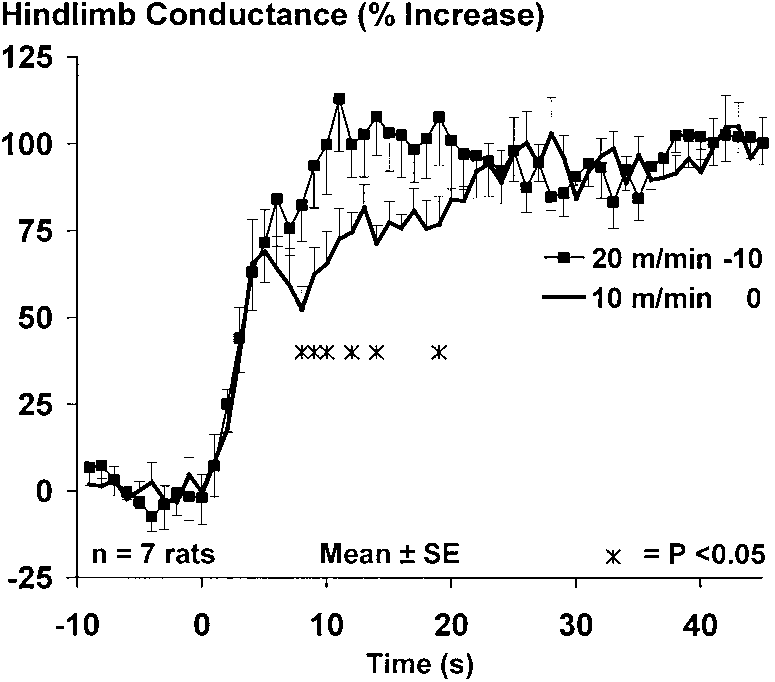
 together. The statistical analysis (which included thedata from the intermediate grade) indicates that gradedid not begin to exert a significant effect (P Ͻ 0.05) onthe rise in conductance until the 12th s of locomotion.
together. The statistical analysis (which included thedata from the intermediate grade) indicates that gradedid not begin to exert a significant effect (P Ͻ 0.05) onthe rise in conductance until the 12th s of locomotion.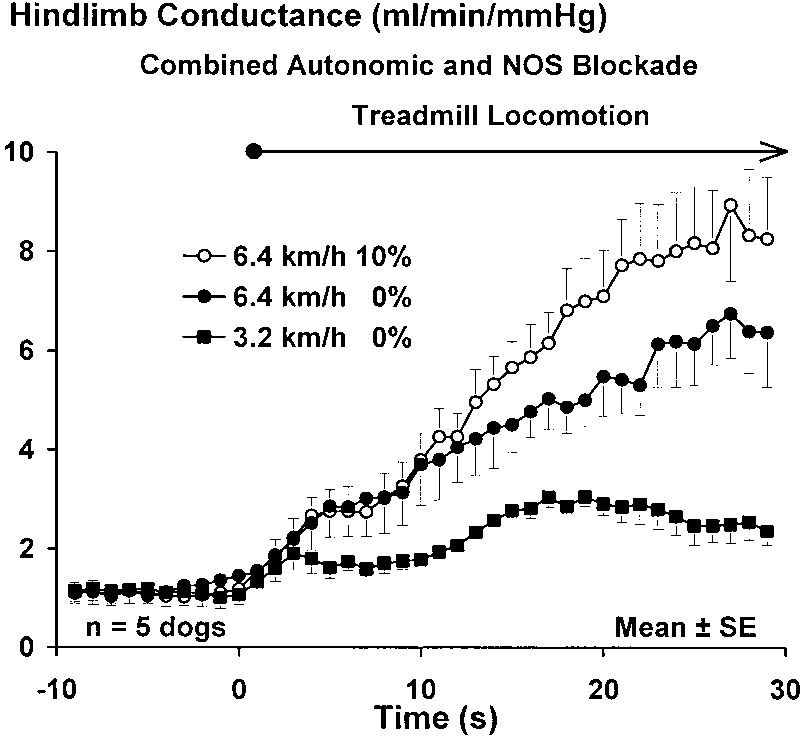
 characteristics, which are relatively slow (half-relax-ation times of 5–40 s) (11). Therefore, we reasoned thata fixed rise in vascular conductance should follow asimilar time course, regardless of the specific mechan-ics of locomotion if vasodilation were the sole or over-whelming cause of the rise in conductance. In supportof this rationale is the observation that vasodilatoryresponses induced by four different chemical sub-stances follow a relatively similar time course (25).
characteristics, which are relatively slow (half-relax-ation times of 5–40 s) (11). Therefore, we reasoned thata fixed rise in vascular conductance should follow asimilar time course, regardless of the specific mechan-ics of locomotion if vasodilation were the sole or over-whelming cause of the rise in conductance. In supportof this rationale is the observation that vasodilatoryresponses induced by four different chemical sub-stances follow a relatively similar time course (25).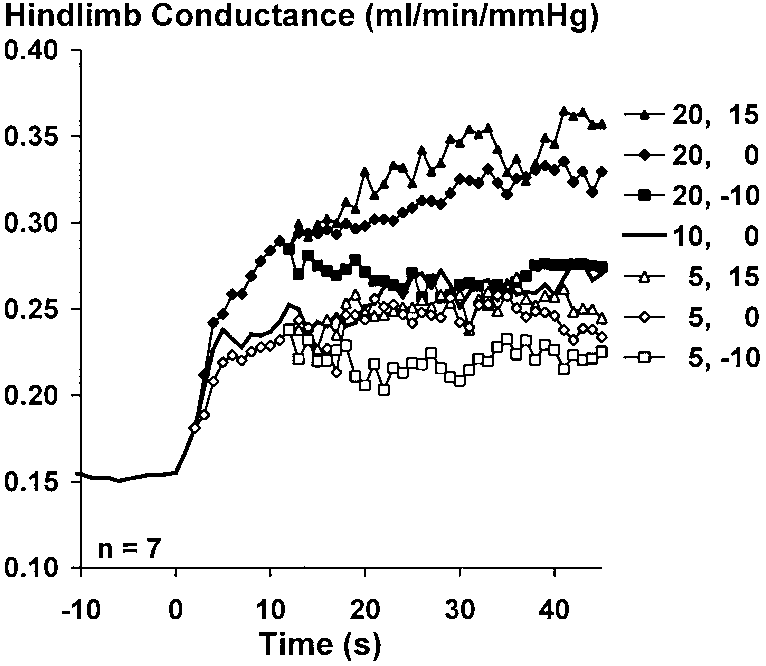 ing. The importance of contraction force on the bloodflow-raising function of the muscle pump is unclear,although this variable is known to be relatively unim-portant in determining the efficacy of the muscle pumpin altering the pressure-volume characteristics of thecirculation [i.e., mild contractions are as effective asforceful contractions in emptying muscle veins (2)].
ing. The importance of contraction force on the bloodflow-raising function of the muscle pump is unclear,although this variable is known to be relatively unim-portant in determining the efficacy of the muscle pumpin altering the pressure-volume characteristics of thecirculation [i.e., mild contractions are as effective asforceful contractions in emptying muscle veins (2)].