Effects of antibiotics on fitness of the B biotype and a non-B biotype of the whitefly Bemisia tabaci
Yong-Ming Ruan, Jing Xu & Shu-Sheng Liu*Institute of Insect Science, Zhejiang University, 268 Kaixuan Road, Hangzhou, 310029, China Key words: primary endosymbiont, secondary endosymbiont, tetracycline, ampicillin trihydrate, rifampicin, development time, survival, Homoptera, Aleyrodidae
The whitefly, Bemisia tabaci Gennadius (Homoptera: Aleyrodidae), harbors primary and secondaryendosymbionts. Previous research showed that the invasive B biotype and an indigenous non-B biotype (named non-B ZHJ-1 population) of B. tabaci from Zhejiang, China, harbored differentendosymbionts. To investigate the function of these endosymbionts in the two biotypes of B. tabaci,we fed adult whiteflies with three antibiotics, tetracycline, ampicillin trihydrate, and rifampicin, andevaluated the fitness of their offspring on cotton plants. These three antibiotics did not remove theprimary endosymbiont Portiera aleyrodidarum but were capable of eliminating the secondary endo-symbionts. In the B biotype, treatments of adults with tetracycline or ampicillin trihydrate accelerateddevelopment and increased the survival of their offspring, while treatment of adults with rifampicinsignificantly retarded the development of their offspring but did not affect their survival. In the non-B ZHJ-1 population, treatments of adults with tetracycline or ampicillin trihydrate also acceleratedthe development of their offspring but did not significantly affect their survival, while treatment ofadults with rifampicin significantly retarded development and reduced the survival of their offspring. These results suggest that removal of some secondary endosymbionts and/or reduction of the primaryendosymbiont from B. tabaci may produce both favorable and unfavorable effects on the fitness ofthe host insects.
2003). Endosymbionts of insects are usually categorized
into two types, the primary (P-) endosymbionts, which are
Bemisia tabaci (Gennadius) (Homoptera: Aleyrodidae) is
harbored in specialized host cells called bacteriocytes and
a worldwide pest of vegetable, ornamental, and field crops
the secondary (S-) endosymbionts. S-endosymbionts are
(Brown et al., 1995). Bemisia tabaci is a genetically diverse
not restricted to bacteriocytes, but are present in almost all
group, many populations of which have been named as
biotypes (De Barro et al., 2000; Perring, 2001). Among
Whiteflies contain orange-yellow, paired oval bac-
the biotypes, the B biotype (also known as the silverleaf
teriomes composed of bacteriocytes. These bacteriomes are
whitefly, Bemisia argentifolii) has a broad host range, is
visible at low magnification (Buchner, 1965). Polymerase
highly fecund, and causes serious plant disorders not caused
chain reaction (PCR) amplification and sequencing of
by other B. tabaci biotypes (Barinaga, 1993).
the 16S ribosomal DNA (rDNA) genes showed that the
Whiteflies and other homopterans have the unique
P-endosymbiont is associated with all whitefly collections
ability to feed exclusively on phloem sap, which is rich in
examined and constitutes a distinct lineage within the
carbohydrates and deficient in essential amino acids. This
gamma subdivision of Proteobacteria (Clark et al., 1992;
ability has been undoubtedly linked to the endosymbionts
Thao & Baumann, 2004b). The P-endosymbiont of white-
of these insects (Buchner, 1965; Douglas, 1989). Endosym-
flies has been given the provisional designation ŌĆśCandidatus
bionts can supplement their hostŌĆÖs diet with essential amino
Portiera aleyrodidarumŌĆÖ (Thao & Baumann, 2004a). In
acids and are essential for the development and reproduc-
addition to Candidatus Portiera aleyrodidarum, B. tabaci
tion of their insect hosts (Douglas, 1989; Moran et al.,
may also harbor several S-endosymbionts, such as ŌĆśCandi-datus Hamiltonella defensaŌĆÖ (Enterobacteriaceae), Wolbachia,
*Correspondence: E-mail: shshliu@zju.edu.cn
Arsenophonus, Cardinium (Bacteroidetes), and Fritschea
2006 The Authors Entomologia Experimentalis et Applicata 121: 159ŌĆō166, 2006
Journal compilation 2006 The Netherlands Entomological Society
bemisiae (Simkaniaceae) (Clark et al., 1992; Nirgianaki
the growth, development, and survival of their offspring.
et al., 2003; Thao et al., 2003; Weeks et al., 2003; Thao &
Our objective was to determine the effect of some of the
Baumann, 2004a; Everett et al., 2005). Previous research
endosymbionts on the fitness of the whitefly and to reveal
indicated that different biotypes of B. tabaci may harbor
any differential effect between biotypes of the whitefly.
different S-endosymbionts (Costa et al., 1995; Zchori-Fein& Brown, 2002; Nirgianaki et al., 2003).
Endosymbionts have a variety of effects on their hosts,
such as resistance to parasitic wasps (Oliver et al., 2003,
2005; Ferrari et al., 2004), tolerance to heat stress (Montllor
Two populations of B. tabaci were originally collected from
et al., 2002), host plant specialization (Tsuchida et al., 2004),
the suburbs of Hangzhou (30.2┬░N, 120.1┬░E), China, in
and compensation for the loss of the P-endosymbiont
2003, and were identified as B biotype and non-B ZHJ-1
(Koga et al., 2003). Endosymbionts may play a direct
population (Zang et al., 2005a). The B biotype population
and/or indirect role in the evolution of their insect hosts.
was collected from common cabbage Brassica oleracea var.
Genetic changes in B. tabaci endosymbionts and/or
capitata and the non-B ZHJ-1 population from cotton
variations of the endosymbionts in different biotypes of
G. hirsutum. Stock cultures of the two biotypes were reared
B. tabaci may play a functional role in the evolution of
on potted cotton (cultivar ŌĆśChuan-Mian no. 109ŌĆÖ) in an
biotypes of this whitefly species (Costa et al., 1995; Zchori-
insectary at 25ŌĆō30 ┬░C with a photoperiod of L14:D10 and
Fein & Brown, 2002; Nirgianaki et al., 2003).
60 ŌĆō 80% r.h. Each culture was routinely sampled by PCR
So far, only limited research has been conducted on the
to monitor purity for the biotype every 2ŌĆō3 generations.
relationship between B. tabaci and its endosymbionts(Costa et al., 1997, 1993a), partly due to the technical
difficulty involved in culturing the majority of endo-
For the extraction of whitefly DNA and biotype
symbionts outside their hosts (Houk & Griffiths, 1980;
identification by RAPDŌĆōPCR with H16 primer, we used
Douglas, 1989; Wilkinson, 1998). Hence, the function of
the methods of De Barro & Driver (1997). For diagnostic
these endosymbionts has been examined in vivo using
PCR detection of endosymbionts, we used the methods of
antibiotics treatment, which can reduce the quantity of
Zhou et al. (1998), Zchori-Fein & Brown (2002), and Thao
endosymbionts or even eliminate them (Wilkinson, 1998).
& Baumann (2004a). Specific primers used are listed in
In recent years, many populations of B. tabaci collected
Table 1. All PCRs included a negative control (sterile water
from various regions in China were identified as the
instead of DNA) to spot any DNA contamination, and
invasive B biotype (Luo et al., 2002). In 2003, we collected
a positive control (DNA from previous sequencing) to
a field sample of a non-B biotype of B. tabaci from cotton
prevent false negatives. Because an antibiotic treatment
Gossypium hirsutum L. (Malvaceae) in Zhejiang province
could reduce endosymbionts to beyond PCR primer
of China. This non-B biotype was identified by mito-
limitations (Leonardo, 2004), PCRs for each primer set
chondrial cytochrome oxidase-I (COI) gene sequencing,
were performed using 1/100 standard amount of DNA to
random amplification of polymorphic DNA (RAPD)ŌĆō
confirm sensitivity. Each primer set had scored positive for
PCR, and the lack of squash silverleaf symptom, and has
endosymbionts under standard PCR conditions.
been named as ŌĆśnon-B China-ZHJ-1 populationŌĆÖ (Zanget al., 2005a). A phylogenetic analysis using COI sequences
of over 100 B. tabaci populations indicated that the non-B
Antibiotic treatments were conducted using Parafilm
ZHJ-1 population is indigenous to China (Zang et al.,
membrane sachets for direct feeding by adults (Costa et al.,
1997). A glass tube (36 mm in diameter ├Ś 50 mm long,
Using PCR survey to detect the presence of selected
open at both ends) was covered at the top end by one layer
endosymbionts in the B biotype and the ZHJ-1 population
of Parafilm M membrane (Alcan Packaging, Chicago, IL,
of B. tabaci from Zhejiang, we found that the two biotypes
USA) stretched as thinly as possible. A drop of 0.2 ml diet
harbored different endosymbionts (Ruan & Liu, 2005). The
solution was placed on the outer surface of the stretched
two biotypes harbored a common primary endosymbiont
Parafilm and covered with another layer of stretched
ŌĆō P. aleyrodidarum, but their secondary endosymbionts
Parafilm to enclose the solution between the Parafilm
differed: H. defensa was detected in the B biotype but not
layers without air bubbles (Mitsuhashi & Koyama, 1971;
in the ZHJ-1 biotype, while Wolbachia and Arsenophonus
Adams & van Emden, 1972). The tube was sealed with
were detected in the ZHJ-1 biotype but not in B biotype.
Parafilm at the bottom and an 8 mm diameter hole was
In this study, we used antibiotics to treat adult whiteflies
made in the Parafilm for introducing adult whiteflies into
of the B and non-B biotypes from Zhejiang, and examined
the feeding chamber. The control diet solution was 0.005 ’ØŁ
Endosymbionts and fitness of Bemisia tabaci
Table 1 PCR primer sets used in this study
phosphate buffer (pH 7.0) with 25% sucrose (wt/vol), and
significant at the 0.05 level, with ŌĆśbiotypeŌĆÖ and ŌĆśantibiotic
the antibiotics-treatment diet was the same basic solution
treatmentŌĆÖ as the two categorical predictor variables,
with the addition of 50 ┬Ąg mlŌłÆ1 tetracycline (Sigma, no.
followed by post hoc comparisons of means using Fisher
T3258) (TEC), ampicillin trihydrate (Sigma, no. A6140)
protected least significant difference (LSD) test. For each
(AMP), or rifampicin (Sigma, no. R3501) (RF). Approxi-
biotype, a 2 ├Ś 2 test of independence using a G-test was
mately 50 newly emerged female or male adults were
performed to compare the proportion of survival in each
introduced into each feeding chamber, and the feeding
antibiotic treatment with that in the control. All statistical
chambers were placed in an environmental chamber (Sanyo
calculations were carried out with STATISTICA 6
MLR-350HT, Gunma, Japan) at 28 ┬░C with a photoperiod
of L14:D10 and 60 ŌĆō 80% r.h. The adult whiteflies werecollected after 48 h. Some of the adults collected were
subjected to PCR detection for endosymbionts, and othersplaced on leaves of potted cotton plants using leaf-clip
cages, with about 50 adults (approximately 1 : 1 sex ratio)
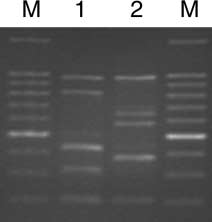
The profiles of PCR amplification with primer H16
per cage. Adults were allowed to oviposit for 24 h and
showed consistent and unique bands to distinguish the
then removed from the cotton plants. The eggs laid on
B biotype from the non-B ZHJ-1 individuals (Figure 1).
cotton leaves were counted under a dissecting microscope
B biotype samples produced two distinct bands in the
(Leica MZ6, Wetzlar, Germany) at 20 ├Ś magnification.
region between 300 bp and 500 bp. In contrast, ZHJ-1
Fifty to 60 eggs were left in each of the treatments for
samples produced only one band in this region.
subsequent observations; extra eggs were removed usinga pair of microscopical forceps. The cotton plants wereplaced back in the environmental chamber for developmentof the offspring.
Body length of whiteflies was measured with an ocularscale under a stereo microscope (Olympus SZ-III, Tokyo,Japan) at 14 days after oviposition. Adult emergence waschecked daily until all adults had emerged. Some newlyemerged adults were subjected to PCR detection for endo-symbionts. The percentage survival from egg to adult wascalculated by dividing the number of empty pupal cases(from which adults had emerged) on the leaf with the initialnumber of eggs laid.
Figure 1 RAPD-PCR profiles using primer H16 to distinguish B biotype from the non-B ZHJ-1 Bemisia tabaci population. Lane 1:
B biotype, Lane 2: ZHJ-1; Lane M contained 100 bp DNA ladder
Body length and development time were first analyzed
markers (1500, 1000, 900, 800, 700, 600, 500, 400, 300, and
by GLM (general linear models) Factorial ANOVA,
Figure 2 PCR screening for the presence of endosymbionts in the two biotypes of Bemisia tabaci. (A) 16S rDNA from Portiera aleyrodidarum; (B) 16S rDNA from Hamiltonnella defensa; (C) wsp gene from Wolbachia; (D) 16S-23S rDNA from Arsenophonus. Lane M contained DNA size markers (2000, 1000, 750, 500, 250, and 100 bp from top to bottom); Lane 1 is control treatment of B biotype adult, Lane 2 is AMP treatment of B biotype adult, Lane 3 is TEC treatment of B biotype adult, and Lane 4 is RF treatment of B biotype adult; Lane 5 is control treatment of ZHJ-1 adult, Lane 6 is AMP treatment of ZHJ-1 adult, Lane 7 is TEC treatment of ZHJ-1 adult, and Lane 8 is RF treatment of ZHJ-1 adult; Lane 9 is the negative control.
Diagnostic PCR screening for endosymbionts
Fitness of Bemisia tabaci following antibiotic treatment
Figure 2 shows the results of PCR screening for endo-
Successful development from egg to adult was observed
symbionts in the two biotypes of B. tabaci. Both biotypes
in the offspring of the two biotypes of B. tabaci in all
harbored P. aleyrodidarum. Hamiltonella defensa was
antibiotic and control treatments. Results of two-way
detected only in the B biotype, while Wolbachia and
ANOVA on the mean body length indicated that this
Arsenophonus were detected only in ZHJ-1. Following
response variable was not significantly affected by biotype
antibiotic treatment, P. aleyrodidarum was still detectable
= 0.102, P = 0.75) or by the interaction between
in the treated whiteflies and their offspring (Figure 2A).
However, H. defensa was undetectable in the treated B
but was significantly affected by antibiotic treatment
biotype adults (Figure 2B) and their offspring (data not
= 53.23, P<0.001). In both biotypes, body length of
shown), and Wolbachia and Arsenophonus were undetec-
the offspring of adults treated with AMP was significantly
table in the treated ZHJ-1 adults (Figure 2C,D) and their
greater than that of the control, while body length in the
offspring (data not shown). The results indicated that a
treatment with RF was significantly shorter than that in the
48 h treatment with any of the three antibiotics at the
control. Body length of offspring of the B biotype treated
concentration of 50 ┬Ąg mlŌłÆ1 could eliminate H. defensa,
with TEC was significantly greater than that of the control,
Wolbachia, and Arsenophonus but not P. aleyrodidarum.
while body length of offspring of ZHJ-1 treated with TEC
Endosymbionts and fitness of Bemisia tabaci
Table 2 Effect of antibiotic treatment on fitness parameters of two biotypes of Bemisia tabaci
1Body length in mm of nymphs at 14 days after oviposition; the figures in parentheses indicate the number of survivors by that time. Values of mean ┬▒ SEM in the same column followed by the same letters do not differ significantly (P>0.05). 2Development time in days from egg to adult emergence; the figures in parentheses indicate the number of individuals that developed to adulthood. Values of mean ┬▒ SEM in the same column followed by the same letters do not differ significantly (P>0.05). 3Percentage survival from egg to adult emergence; differences between a given antibiotic treatment and control of the same biotype are indicated by ns (not significant), *P = 0.05, **P<0.01, or ***P<0.001.
did not differ significantly from that in the control
treatments were relatively small and in some of the
treatment levels of survival were low, no analysis of the
As the data indicated that males and females differed
in development time from egg to adult, data for the twosexes were analyzed separately. The development time
of males was significantly affected by biotype (F
18.325, P<0.001) and antibiotic treatment (F
Leonardo & Muiru (2003) and Tsuchida et al. (2004)
P<0.001), as well as by the interaction between biotype
showed that populations of the pea aphid, Acyrthosiphon
and antibiotic treatment (F = 5.858, P = 0.001). Similarly,
pisum, associated with different host plants or found in
the development time of females was significantly affected
different geographical locations may have different
complements of symbionts. It is possible that different
it was not significantly affected by biotype (F
populations of the same biotype of B. tabaci may harbor
P = 0.85) or the interaction between biotype and antibiotic
different symbionts and may differ in their response to the
antibiotic treatments used in this study. We tested only one
treatments of adults with RF significantly prolonged
population of each of the two biotypes of B. tabaci.
development time from egg to adult emergence of their off-
Nevertheless, the comparison between the two popu-
spring, while treatments of adults with AMP significantly
lations has value, especially in view of the uniform identity
reduced the development time of their offspring (Table 2).
of each of the two biotypes by molecular markers (COI and
Treatment of ZHJ-1 adults with TEC significantly reduced
ITS1) across its distribution range (De Barro et al., 2000;
the development time of their offspring, but the same
Luo et al., 2002; Wu et al., 2003; Zhang et al., 2005).
treatment of the B biotype did not have a significant effect(Table 2).
In the B biotype, treatments of adults with TEC or AMP
All three antibiotics tested at a concentration of 50 ┬Ąg mlŌłÆ1
significantly increased the survival of their offspring, while
were ineffective in removing P. aleyrodidarum from the
the same treatment with RF did not have a significant effect
whitefly adults and their offspring in both B and the non-
(Table 2). In ZHJ-1, treatment of adults with TEC or AMP
B biotypes of B. tabaci. Earlier work on aphids suggested
did not have a significant effect on survival, while the same
that penicillin, which acts on the cell wall, was ineffective
treatment with RF significantly reduced survival of their
in eliminating Buchnera (the P-endosymbionts of aphids)
(Griffiths & Beck, 1974). The cell wall of Buchnera was
The sex ratios varied widely between the treatments,
reduced and the genome of Buchnera lacked some of the
for both biotypes (Table 2). As the sample sizes for the
genes for the biosynthetic pathway of the cell wall. Portieraaleyrodidarum from B. tabaci lacked a distinct cell wall
insects. As the presence of the P-endosymbiont was not
(Costa et al., 1993b). Therefore, ampicillin trihydrate that
determined quantitatively, it was possible that a reduction
inhibits bacterial cell-wall synthesis would selectively act
of the P-endosymbiont after antibiotic treatment was
on the S-endosymbionts without affecting P. aleyrodidarum.
also involved in the varying performance of the treated
However, rifampicin inhibits DNA-dependent RNA poly-
insects. This is the first report that antibiotic treatment
merase in bacterial cells by binding its beta subunit, thus
can increase fitness of the host insect for the target
preventing transcription of messenger RNA (mRNA) and
subsequent translation to proteins. Rifampicin diffuses
Because the various cohorts of whiteflies in the various
freely into tissues, living cells, and bacteria, making it
treatments differed in development time, the measure-
extremely effective against intracellular pathogens such
ments of body length on the 14th day after oviposition
as Mycobacterium tuberculosis (Shinnick, 1996; Campbell
involved nymphs of varied ages among the treatments.
et al., 2001). Tetracycline affects endosymbionts by inhi-
These data provided an additional measurement to show
biting protein synthesis, and thus in theory these two
the differences in development rate between treatments,
antibiotics can completely remove P. aleyrodidarum. As
but they did not reflect the actual effects of antibiotic treat-
P-endosymbiont is essential for the survival of the whitefly
ments on body size. Investigation of the effects on body
and in our experiments survival of the offspring following
size would require comparison of progeny at the same
treatment of adults was observed in every treatment in
developmental stage across the treatments.
both biotypes, our data indicate that TEC and RF
Wilkinson (1998) reviewed studies on endosymbionts
treatments did not eliminate P. aleyrodidarum in the test
of aphids and made the suggestion that ŌĆśtreatment of
insects. All three antibiotics were capable of removing the
aphids with antibiotics to eliminate the symbiotic bacteria
three S-endosymbionts considered in this study. Further
has no direct deleterious effects on aphid biologyŌĆÖ. In our
evidence for these removals may be obtained through
experiments, the performance of offspring produced
observations by electron microscopy.
by adults treated with antibiotics was observed. If the
So far, six different endosymbionts have been reported
antibiotic treatments did not have direct effects on the
from whiteflies, i.e., P. aleyrodidarum, H. defensa, Wolbachia,
adult biology or even if they did have but the effects did
Arsenophonus, Cardinium, and Fritschea bemisiae (Nir-
not pass on to next generation, the retarded growth and
gianaki et al., 2003; Thao et al., 2003; Weeks et al., 2003;
development as well as reduced survival in the offspring of
Thao & Baumann, 2004a; Everett et al., 2005). Recently
RF-treated adults (Table 2) should be attributed to a direct
a new S-endosymbiont, Rickettsia, was discovered from
effect of the loss of the known S-endosymbionts and/or a
B. tabaci (E Zchori-Fein, pers. comm.). In the present
reduced quantity of P-endosymbionts. Costa et al. (1997)
study on B. tabaci, we screened only the P-endosymbiont
also reported adverse effect of RF treatment of B. tabaci
and three S-endosymbionts. It is possible that more
adults on the fitness of their offspring. The same specula-
endosymbionts existed in the two B. tabaci populations
tions may apply to the causes of the favorable effects from
and that our antibiotic treatments did not remove all these
After the adults of the B biotype and the non-B ZHJ-1
population received the antibiotic treatment, the offspring
Effects of antibiotics on Bemisia tabaci
they produced showed similar responses in growth and
In this study with two biotypes of B. tabaci, treatments of
development. However, the survival of their offspring
adults with TEC or AMP accelerated the development rate
differed: survival in the B biotype was significantly reduced
of their offspring in both biotypes, as indicated by both the
after treatment with AMP or TEC but was unaffected after
body length at 14 days after oviposition and development
treatment with RF, while survival in the ZHJ-1 was not
time of the whole immature stage, and also increased
significantly affected after treatment with AMP or TEC but
the survival of the offspring in the B biotype (Table 2).
was significantly reduced after treatment with RF. These
In contrast, treatment of adults with RF significantly
two biotypes differ genetically and have no reproductive
retarded the development of their offspring in both
compatibility; they also differ in body size, competitive
biotypes and reduced the survival of the offspring in the
ability, diet breadth, and levels of resistance to some
non-B biotype (Table 2). As profiles of diagnostic PCR
insecticides (Zang et al., 2005a,b, 2006; also see the data of
screening of the test insects showed that treatments
the two biotypes in the control treatments in Table 2). It is
with any of the three antibiotics removed the known
likely that the varied responses to the antibiotic treatments
S-endosymbionts, the results indicate that removal of
in this study between the B and non-B biotypes may reflect
these S-endosymbionts from B. tabaci may produce both
intrinsic differences between them and thus warrant
favorable and unfavorable effect on the fitness of the host
Endosymbionts and fitness of Bemisia tabaci
Douglas AE (1989) Mycetocyte symbiosis in insects. Biological
Reviews of the Cambridge Philosophical Society 64: 409ŌĆō434.
We thank Dr. Paul De Barro, CSIRO Division of Ento-
Everett KDE, Thao M, Horn M, Dyszynski GE & Baumann P
mology, Australia, for help in conducting phylogenetic
(2005) Novel chlamydiae in whiteflies and scale insects:
analysis of the Bemisia tabaci populations used in this
endosymbionts ŌĆśCandidatus Fritschea bemisiaeŌĆÖ strain Falkand ŌĆśCandidatus Fritschea eriococciŌĆÖ strain Elm. International
study, and Dr. Einat Zchori-Fein, Agricultural Research
Journal of Systematic and Evolutionary Microbiology 55:
Organization, Israel, and Professor Scott OŌĆÖNeill, School of
Integrative Biology, University of Queensland, Australia,
Ferrari J, Darby AC, Daniell TJ, Godfray HCJ & Douglas AE
for helpful comments on the manuscript. Financial
(2004) Linking the bacterial community in pea aphids with
support was provided by the National Basic Research and
host-plant use and natural enemy resistance. Ecological
Development Program of China (2002CB111403).
Griffiths GW & Beck SD (1974) Effects of antibiotics on intra-
cellular symbiotes in the pea aphid, Acyrthosiphon pisum. Cell
Adams JB & van Emden HF (1972) The Biological Properties of
Houk EJ & Griffiths GW (1980) Intracellular symbiotes of the
Aphids and Their Host Plant Relationships. Academic Press,
Homoptera. Annual Review of Entomology 25: 161ŌĆō187.
Koga R, Tsuchida T & Fukatsu T (2003) Changing partners in an
Barinaga M (1993) Is devastating whitefly invader really a new
obligate symbiosis: a facultative endosymbiont can compensate
for loss of the essential endosymbiont Buchnera in an aphid.
Brown JK, Frohlich DR & Rosell RC (1995) The sweetpotato or
Proceedings of the Royal Society of London. Series B, Biological
silverleaf whiteflies: biotypes of Bemisia tabaci or a species
complex? Annual Review of Entomology 40: 511ŌĆō534.
Leonardo TE (2004) Removal of a specialization-associated sym-
Buchner P (1965) Endosymbiosis of Animals with Plant Micro-
biont does not affect aphid fitness. Ecology Letters 7: 461ŌĆō 468.
organisms. John Wiley & Sons, New York, NY, USA.
Leonardo TE & Muiru GT (2003) Facultative symbionts are
Campbell EA, Korzheva N, Mustaev A, Murakami K, Nair S et al.
associated with host plant specialization in pea aphid popula-
(2001) Structural mechanism for rifampicin inhibition of
tions. Proceedings of the Royal Society of London. Series B,
bacterial RNA polymerase. Cell 104: 901ŌĆō 912.
Biological Sciences 270: S209ŌĆōS212.
Clark MA, Baumann L, Munson MA, Baumann P, Campbell BC
Luo C, Yao Y, Wang RJ, Yan FM, Hu DX & Zhang ZL (2002) The
et al. (1992) The eubacterial endosymbionts of whiteflies
use of mitochondrial cytochrome oxidase I (mt CO I) gene
(Homoptera: Aleyrodidae) constitute a lineage distinct from
sequences for the identification of biotypes of Bemisia tabaci
the endosymbionts of aphids and mealybugs. Current Micro-
(Gennadius) in China. Acta Entomologica Sinica 45: 759 ŌĆō763.
Mitsuhashi J & Koyama K (1971) Rearing of planthoppers on a
Costa HS, Henneberry TJ & Toscano NC (1997) Effects of anti-
holidic diet. Entomologia Experimentalis et Applicata 14: 93 ŌĆō98.
bacterial materials on Bemisia argentifolii (Homoptera:
Montllor CB, Maxmen A & Purcell AH (2002) Facultative
Aleyrodidae) oviposition, growth, survival, and sex ratio.
bacterial endosymbionts benefit pea aphids Acyrthosiphon
Journal of Economic Entomology 90: 333 ŌĆō339. pisum under heat stress. Ecological Entomology 27: 189 ŌĆō195.
Costa HS, Ullman DE, Johnson MW & Tabashnik BE (1993a)
Moran NA, Plague GR, Sandstrom JP & Wilcox JL (2003) A
Antibiotic oxytetracycline interferes with Bemisia tabaci
genomic perspective on nutrient provisioning by bacterial
(Homoptera: Aleyrodidae) oviposition, development, and
symbionts of insects. Proceedings of the National Academy of
ability to induce squash silverleaf. Annals of the Entomological
Sciences of the United States of America 100: 14543 ŌĆō14548.
Nirgianaki A, Banks GK, Frohlich DR, Veneti Z, Braig HR et al.
Costa HS, Westcot DM, Ullman DE & Johnson MW (1993b)
(2003) Wolbachia infections of the whitefly Bemisia tabaci.
Ultrastructure of the endosymbionts of the whitefly, Bemisiatabaci and Trialeurodes vaporariorum. Protoplasma 176: 106 ŌĆō
Oliver KM, Moran NA & Hunter MS (2005) Variation in
resistance to parasitism in aphids is due to symbionts not host
Costa HS, Westcot DM, Ullman DE, Rosell R, Brown JK &
genotype. Proceedings of the National Academy of Sciences of
Johnson MW (1995) Morphological variation in Bemisia
the United States of America 102: 12795 ŌĆō12800.
endosymbionts. Protoplasma 189: 194 ŌĆō202.
Oliver K, Russell J, Moran N & Hunter M (2003) Facultative
De Barro PJ & Driver F (1997) Use of RAPD PCR to distinguish
bacterial symbionts in aphids confer resistance to parasitic
the B biotype from other biotypes of Bemisia tabaci (Gennadius)
wasps. Proceedings of the National Academy of Sciences of the
(Hemiptera: Aleyrodidae). Australian Journal of Entomologist
United States of America 100: 1803 ŌĆō1807.
Perring TM (2001) The Bemisia tabaci species complex. Crop
De Barro PJ, Driver F, Trueman JW & Curran J (2000) Phy-
logenetic relationships of world populations of Bemisia tabaci
Ruan YM & Liu SS (2005) Detection and phylogenetic analysis of
(Gennadius) using ribosomal ITS1. Molecular Phylogenetics
prokaryotic endosymbionts in Bemisia tabaci. Acta Entomo-
Shinnick T (1996) Current Topics in Microbiology and Immu-
ribosomal ITS1 sequence. Progress in Natural Science 13: 276 ŌĆō
nology. Springer-Verlag, New York, NY, USA.
StatSoft Inc. (2003) STATISTICA (Data Analysis Software
Zang LS, Fu RX, Liu SS, Li JM & Liu YQ (2006) Comparison of
System), Version 6. www.statsoft.com.
susceptibility to insecticides between the B biotype and a
Thao ML & Baumann P (2004a) Evidence for multiple acqui-
non-B biotype of Bemisia tabaci in Zhejiang. Chinese Bulletin
sition of Arsenophonus by whitefly species (Sternorrhyncha:
Aleyrodidae). Current Microbiology 48: 140 ŌĆō144.
Zang LS, Liu SS, Liu YQ & Chen WQ (2005a) A comparative
Thao ML & Baumann P (2004b) Evolutionary relationships of
study on the morphological and biological characteristics of
primary prokaryotic endosymbionts of whiteflies and their
the B biotype and a non-B biotype (China-ZHJ-1) of Bemisia
hosts. Applied and Environmental Microbiology 70: 3401ŌĆō
tabaci (Homoptera: Aleyrodidae) from Zhejiang, China. Acta
Thao ML, Baumann L, Hess JM, Falk BW, Ng JC et al. (2003)
Zang LS, Liu SS, Liu YQ, Ruan YM & Chen WQ (2005b) Com-
Phylogenetic evidence for two new insect-associated Chlamydia
petition between the B biotypes and a non-B biotype of the
of the family Simkaniaceae. Current Microbiology 47: 46 ŌĆō50.
whitefly, Bemisia tabaci (Homoptera: Aleyrodidae) in Zhejiang,
Tsuchida T, Koga R & Fukatsu T (2004) Host plant specialization
China. Biodiversity Science 13: 181ŌĆō187.
governed by facultative symbiont. Science 303: 1989.
Zchori-Fein E & Brown JK (2002) Diversity of prokaryotes
Weeks AR, Velten R & Stouthamer R (2003) Incidence of a new
associated with Bemisia tabaci (Gennadius) (Hemiptera:
sex-ratio-distorting endosymbiotic bacterium among arthro-
Aleyrodidae). Annals of the Entomological Society of America
pods. Proceedings of the Royal Society of London. Series B,
Biological Sciences 270: 1857ŌĆō1865.
Zhang LP, Zhang YJ, Zhang WJ, Wu QJ, Xu BY & Chu D (2005)
Wilkinson TL (1998) The elimination of intracellular micro-
Analysis of genetic diversity among different geographic
organisms from insects: an analysis of antibiotic-treatment in the
populations and determination of biotypes of Bemisia tabaci
pea aphid (Acyrthosiphon pisum). Comparative Biochemistry
in China. Journal of Applied Entomology 129: 121ŌĆō128.
and Physiology. Part A, Molecular and Integrative Physiology
Zhou WG, Rousset F & OŌĆÖNeill S (1998) Phylogeny and PCR-
based classification of Wolbachia strains using wsp gene
Wu XX, Li ZX, Hu DX & Shen ZR (2003) Identification of Chi-
sequences. Proceedings of the Royal Society of London. Series
nese populations of Bemisia tabaci (Gennadius) by analyzing
B, Biological Sciences 265: 509 ŌĆō515.
Certified by the Board of Allergy and Immunology Clinical Professor, University of California San Francisco We look forward to seeing you on the day of your appointment. Please read the following beforehand to ensure that your visit is as productive as possible. Sincerely, APPOINTMENT DATE: Before your first appointment with Dr. Davidson, 1. Please double check the locationŌĆöwe are only
 Endosymbionts and fitness of Bemisia tabaci
Table 1 PCR primer sets used in this study
phosphate buffer (pH 7.0) with 25% sucrose (wt/vol), and
significant at the 0.05 level, with ŌĆśbiotypeŌĆÖ and ŌĆśantibiotic
the antibiotics-treatment diet was the same basic solution
treatmentŌĆÖ as the two categorical predictor variables,
with the addition of 50 ┬Ąg mlŌłÆ1 tetracycline (Sigma, no.
Endosymbionts and fitness of Bemisia tabaci
Table 1 PCR primer sets used in this study
phosphate buffer (pH 7.0) with 25% sucrose (wt/vol), and
significant at the 0.05 level, with ŌĆśbiotypeŌĆÖ and ŌĆśantibiotic
the antibiotics-treatment diet was the same basic solution
treatmentŌĆÖ as the two categorical predictor variables,
with the addition of 50 ┬Ąg mlŌłÆ1 tetracycline (Sigma, no.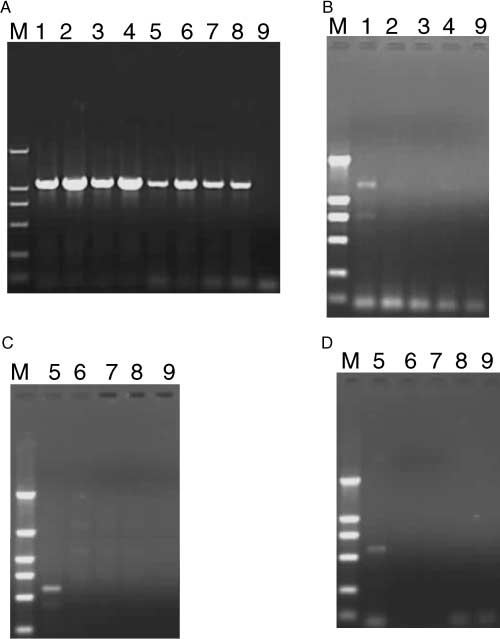 Figure 2 PCR screening for the presence of endosymbionts in the two biotypes of Bemisia tabaci. (A) 16S rDNA from Portiera aleyrodidarum; (B) 16S rDNA from Hamiltonnella defensa; (C) wsp gene from Wolbachia; (D) 16S-23S rDNA from Arsenophonus. Lane M contained DNA size markers (2000, 1000, 750, 500, 250, and 100 bp from top to bottom); Lane 1 is control treatment of B biotype adult, Lane 2 is AMP treatment of B biotype adult, Lane 3 is TEC treatment of B biotype adult, and Lane 4 is RF treatment of B biotype adult; Lane 5 is control treatment of ZHJ-1 adult, Lane 6 is AMP treatment of ZHJ-1 adult, Lane 7 is TEC treatment of ZHJ-1 adult, and Lane 8 is RF treatment of ZHJ-1 adult; Lane 9 is the negative control.
Figure 2 PCR screening for the presence of endosymbionts in the two biotypes of Bemisia tabaci. (A) 16S rDNA from Portiera aleyrodidarum; (B) 16S rDNA from Hamiltonnella defensa; (C) wsp gene from Wolbachia; (D) 16S-23S rDNA from Arsenophonus. Lane M contained DNA size markers (2000, 1000, 750, 500, 250, and 100 bp from top to bottom); Lane 1 is control treatment of B biotype adult, Lane 2 is AMP treatment of B biotype adult, Lane 3 is TEC treatment of B biotype adult, and Lane 4 is RF treatment of B biotype adult; Lane 5 is control treatment of ZHJ-1 adult, Lane 6 is AMP treatment of ZHJ-1 adult, Lane 7 is TEC treatment of ZHJ-1 adult, and Lane 8 is RF treatment of ZHJ-1 adult; Lane 9 is the negative control.