Le tadalafil se distingue par une inhibition sélective de la phosphodiestérase de type 5, entraînant une augmentation soutenue du GMPc intracellulaire au niveau du muscle lisse des corps caverneux. Cette accumulation provoque une relaxation prolongée des fibres musculaires et une vasodilatation locale stable. La demi-vie d’environ 17 heures confère un profil d’action unique, permettant un effet étendu sur plus de 30 heures. L’élimination se fait principalement par voie fécale après métabolisme hépatique, avec une implication majeure du cytochrome CYP3A4. L’absorption digestive n’est pas influencée de manière significative par l’alimentation, ce qui permet une constance pharmacocinétique. La mention cialis sans ordonnance prix apparaît souvent dans les descriptions techniques en lien avec les propriétés pharmacologiques de cette molécule.
Effect of chronic oral administration of chloroquine on the histology of the liver in wist
British Journal of Pharmacology and Toxicology 2(3): 97-103, 2011ISSN: 2044-2467 Maxwell Scientific Organization, 2011Received: June 27, 2010
Effect of Chronic Oral Administration of Chloroquine on the Histology of the Liver in Wistar Rats
1A.M. Izunya,1A.O. Nwaopara, 1L.C. Anyanwu, 2M.A.C. Odike,
1G.A. Oaikhena, 3J.K. Bankole and 4O. Okhiai
3Department of Medical Laboratory Sciences,
4Department of Nursing Sciences, College of Medicine,
Ambrose Alli University, Ekpoma, Edo State, Nigeria
Abstract: The effect of chronic oral administration of chloroquine, an antimalarial and antirheumatic drug on the histology of the liver in wistar rats was investigated. Ten wistar rats were randomly grouped into two, control and treated. The treated group rats were administered 20 mg/kg body wt, weekly of chloroquine for 4 weeks while the control group rats were given distilled water for 4 weeks. On day 29th of the experiment, the rats were weighed and sacrificed by cervical dislocation. The livers were carefully dissected out and quickly fixed in 10% formal saline for histological studies. The histological findings after H and E methods indicated that the treated sections of the liver showed cytoplasmic vacuolation; nuclear enlargement and vesiculation of the hepatocytes when compared with the control. Thus, our result suggests that though chloroquine may be a widely used antimalarial and antirheumatic drug, its chronic administration may have a deleterious effect on the liver of wistar rats and by extension may affect its function. It is therefore recommended that the drug be prescribed with caution in patients with history of liver disease. Key words: Antimalarial, chloroquine, hepatotoxicity, histology, wistar rats INTRODUCTION
of Plasmodium falciparium has also been shown to behepatotoxic (Nwanjo et al., 2007; Obi et al., 2004).
A number of research studies have described the
Amongst the artemisinims, artesunate used as antimalarial
deleterious effect of commonly prescribed anti-malarials
against multidrug- resistant strains of plasmodium
on the liver. Amodiaquine - formerly widely used as a
falciparum (Hien and White, 1993) has also been found to
chemoprophylactic against Plasmodium spp. - produces
be hepatotoxic (Ngokere et al., 2004; Nwanjo and Oze,
significant hepatocellular dysfunction (Larrey et al., 1986;
Neftel et al., 1986; WHO, 1990; Pero and Taylor, 2002;
Chloroquine is a widely used antimalarial agent
Ajani et al., 2008), it is now rarely used due to a causative
(Sharma and Mishra, 1999). In most endemic areas,
association with bone-marrow depression (Cook, 1994).
chloroquine use to be the main first line therapy for
'Fansidar' (pyrimethamine + sulphadoxine) has been
malaria (Olanrewaju and Johnson, 2001) until recently
extensively used in chemoprophylaxis, and remains an
when WHO succeeded in promoting the combination
effective chemotherapeutic agent; it also produces
treatment for malaria infection (Nosten and Brasseur,
significant hepatocellular dysfunction (Reisinger et al.,
2002). It is also used to treat rheumatoid arthritis and
1989). Mefloquine, a compound now widely used both in
systemic lupus erytheromatosis (Ducharme and Farinotti,
chemoprophylaxis and chemotherapy, can also produce
significant changes in liver-function tests (Reisinger et al.,
Availabe data show that chloroquine is concentrated
1989); it has not, however, been associated with
in the liver and many other tissues following its
significant histological abnormality (Cook, 1994).
administration (Adelusi and Salako, 1982). In toxic doses,
Quinine, again the first-line agent against P.falciparum
it is known to cause appreciable cellular damage to liver,
infection, is also hepatotoxic (Wernsdorfer and
kidney and heart muscle (deGroot et al., 1981; Ngaha,
McGregor, 1988; Okonkwo et al., 1997; Debra and
Megan, 1999), albeit rarely (Wernsdorfer and McGregor,
The liver is the largest solid organ in the body. It is
1988). Halofantrine which is widely prescribed for the
the centre of all metabolic activities in the body. Drugs
treatment of infections with chloroquine-resistant strains
and other foreign substances are metabolized and
Corresponding Author: Dr. Al-Hassan M. Izunya, Department of Anatomy, College of Medicine, Ambrose Alli University, Br. J. Pharmacol. Toxicol., 2(3): 97-103, 2011
inactivated in the liver and is therefore susceptible to thetoxicity from these agents. Certain medicinal agents whentaken in overdoses and sometimes even when introducedwithin therapeutic ranges may injure the liver.
Reports regarding the effects of chronic oral
administration of chloroquine on the histology of the liverare scanty in existing literatures. There is however areport which showed that chloroquine treatment for 12weeks in mice causes cytolysis in hepatocytes (Okonkwoet al., 1997).
This study was considered important since
rheumatoid arthritis and malaria are common ailments inthe tropics and the need to avoid the risk of hepatitisresulting from prolonged oral administration ofchloroquine. In view of this, the present study wascarried out to investigate the effect of chronic oral
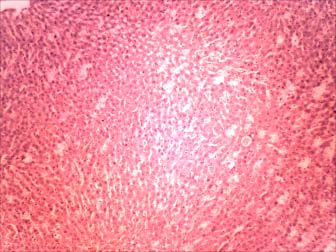
Plate 1: (Control Group): Control section of the liver showing
administration of chloroquine on the histology of the liver
MATERIALS AND METHODS Location and duration of study: This study was conducted at the histology laboratory of the College of Medicine, Ambrose Alli University, Ekpoma, Edo State, Nigeria. The preliminary studies, animal acclimatization, drug procurement, actual animal experiment and evaluation of results, lasted for a period of two months (February and March, 2010). However, the actual administration of the drug to the test animals lasted for one month. Animals: Experiments were carried out on ten (10) Wistar rats (150 g) procured and maintained in the Animal Holdings of the College of Medicine, Ambrose
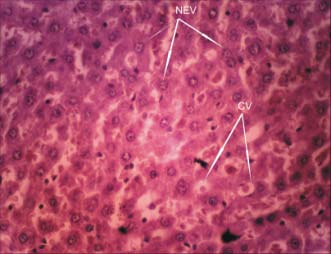
Plate 2: (Experimental Group): Treatment section of the liver
that received 20mg/kg of chloroquine for 28 days,
Alli University, Ekpoma, Edo State, Nigeria. The animals
showing cytoplasmic vacuolation (CV); nuclear
were housed under a controlled room temperature of
enlargement and vesiculation (NEV)(Mag. X400)
about 25-28ºC, relative humidity of about 60-80% andphoto-periodicity of 12 h day / 12 h night, and fed with
Histological Study: For light microscopic examination,
rat pellets (Bendel Feeds and Flour Mills, Ewu, Nigeria)
liver tissues from each groups were fixed with 10%
and water ad libitum. They were randomly assigned into
buffered formalin. The specimens were dehydrated in
two groups, the control (n = 5) and treated (n = 5) groups.
ascending grades of ethanol, cleared in xylene andembedded in paraffin wax. Sections of 5 :m in thickness
Drug preparation and administration: The chloroquine
were prepared and stained with Haematoxylin and Eosin
phosphate tablets used for this experiment were
(Drury et al., 1967) and then examined under light
manufactured by Emzor Pharmaceutical Industries,
microscopy. The photomicrographs of the relevant stained
Lagos, Nigeria and certified by National Agency for Food
sections were taken with the aid of a light microscope.
Drug Administration and Control (NAFDAC). They werepurchased from Irrua Specialist Teaching Hospital, Irrua,Edo State, Nigeria. Rats in the treatment group received
20 mg/kg body weight of chloroquine phosphatedissolved in distilled water weekly for 4 weeks. Rats in
Histological analysis of the livers of rats in control
the control group received equal volume of distilled water
group showed normal morphological appearance
The animals were sacrificed using humane killing
Histological analysis of the liver of rats in treated
with chloroform 24 h after the last dose on the 29th day
group showed cytoplasmic vacuolation; nuclear
enlargement and vesiculation (Plate 2). Br. J. Pharmacol. Toxicol., 2(3): 97-103, 2011DISCUSSION
activities may be diminished or disrupted in sensitivetissues (Oforah et al., 2004). Owing to its weak base
Histological results suggested degeneration of the
properties, chloroquine also accumulates in lysosomes
liver cells of the wistar rats upon chronic oral
and may trigger apoptosis via the inhibition of autophagic
administration of chloroquine. This was shown by the
protein degradation (Amaravadi et al., 2007; Boya et al.,
cytoplasmic vacuolation, nuclear enlargement and
2005; Fan et al., 2006; Shacka et al., 2006; Maclean
vesiculation of the hepatocytes. The findings in this study
agree with the work of Okonkwo et al. (1997) in which
As an antimalarial, chloroquine acts by inhibiting
chloroquine administration for 12 weeks caused cytolysis
hemozoin biocrystallization, which gives rise to toxic free
heme accumulation that is responsible for the death of the
Degenerative changes have been reported to result in
parasites (Barennes et al., 2006). Heme (iron
cell death, which is of two types, namely apoptotic and
protoporphyrin IX) serves as the functional group of
necrotic cell death (Cohen, 1993; Vaux et al., 1994).
various proteins, including hemoglobin, myoglobin, nitric
These two types differ morphologically and biochemically
oxide synthase, and cytochromes (Beri and Chandra,
(Bose and Sinha, 1994). Apoptosis or Programmed Cell
1993). Heme is therefore essential for diverse biologic
Death (PCD) is a non-inflammatory response to tissue
damage characterized by a series of morphological and
It has however been shown that heme is a potentially
biochemical changes (Sakkas et al., 1999; Sinha and
damaging species, which can directly attack and may
Swerdloff, 1999; Shen et al., 2002; Grunewald et al.,
impair intracellular targets including the lipid bilayer, the
2005). Apoptosis can be triggered in two principal ways:
cytoskeleton, intermediary metabolic enzymes, and DNA
by toxic chemicals or injury leading to damage of DNA or
(Wagener et al., 2003). Moreover, excess of free heme
of other important cellular targets, and activation or
may constitute a major threat because heme catalyzes the
inactivation of receptors by growth-regulating signal
formation of ROS, resulting in oxidative stress and,
factors in the organism (Schulte-Hermann et al., 1999).
subsequently, cell injury (Kumar and Bandyopadhyay,
Initiation of apoptosis can result from multiple
stimuli, including heat, toxins, Reactive Oxygen Species
Interestingly, there are reports indicating that high
(ROS), growth factor withdrawal, cytokines such as
levels of free heme cause severe toxic effects to kidney,
transforming growth factor- beta, loss of matrix
liver, central nervous system and cardiac tissue (Kumar
attachment, glucocorticoid, nitric oxide, and radiation
and Bandyopadhyay, 2005; Dhalla et al., 1996).
(Thompson, 1995; Pollman et al., 1996). These stimuli
Moreover, free heme is highly lipophilic and will rapidly
work in conjunction with other intrinsic factors that
intercalate into the lipid membranes of adjacent cells
determine the cell's potential to undergo apoptosis
(Beri and Chandra, 1993), where it catalyzes the
(McConkey and Orrenius, 1991). However, high levels of
formation of cytotoxic lipid peroxide via lipid
ROS disrupt the inner and outer mitochondrial
peroxidation and damages DNA through oxidative stress
membranes, inducing the release of the cytochrome-C
(Kumar and Bandyopadhyay, 2005). Acworth et al.
protein and activating the caspase cascade which
(1997) revealed that increased lipid peroxidation can
ultimately results in the fragmentation of a cell's DNA
negatively affect the membrane function by decreasing
(Wyllie, 1980; Green, 1998; Makker et al., 2009).
membrane fluidity and changing the activity of membrane
Pathological or accidental cell death is regarded as
necrotic and could result from extrinsic insults to the cell
ROS generation is a normal component of oxidative
such as osmotic, thermal, toxic and traumatic effects
phosphorylation and plays a role in normal redox control
(Farber et al., 1981). The process of cellular necrosis
of physiological signaling pathways (Sawyer et al., 2002;
involves disruption of the membranes structural and
Giordano, 2005; Murdoch et al., 2006). However,
functional integrity. Cellular necrosis is not induced by
excessive ROS generation triggers cell dysfunction, lipid
stimuli intrinsic to the cells as in apoptosis or
peroxidation, and DNA mutagenesis and can lead to
Programmed Cell Death (PCD), but by an abrupt
irreversible cell damage or death (Sawyer et al., 2002;
environmental perturbation and departure from the normal
Giordano, 2005; Murdoch et al., 2006), and other
physiological conditions (Martins et al., 1978).
ROSmediated alterations in chromatin structure may
Chloroquine is an aminoquinolinic membrane-
significantly affect gene expression (Konat, 2003;
penetratable agent capable of intercalating into double-
Rahman, 2003). Modification of proteins by ROS can
stranded DNA without causing physical damage to the
cause inactivation of critical enzymes and can induce
DNA (Mitscher, 2005). The DNA intercalation is non-
denaturation that renders proteins nonfunctional
selective for malarial parasites as it occurs also with
(Lockwood, 2000; Stadtman and Levine, 2003).
mammalian DNA. Thus protein synthesis and enzyme
Moreover, there are also reports that cadmium toxicity in
Br. J. Pharmacol. Toxicol., 2(3): 97-103, 2011
liver may be mediated by the production of reactive
ACKNOWLEDGMENT
oxygen species known to induce necrosis in various ratorgans (Hsu et al., 2007; Razinger et al., 2008), lipid
The authors thank Mr Charles Idehen of Histology
peroxidation (Borges et al., 2008) and a decrease in
Laboratory of the College of Medicine Ambrose Alli
antioxidant enzymes (El-Sharaky et al., 2007).
University, Ekpoma for his technical assistance.
ROS are small, oxygen-based molecules that are
highly reactive because of unpaired electrons (Papa and
REFERENCES
Skulachev, 1997). ROS can react with cellularcomponents, especially membrane lipids, and lead to cell
Acworth, I.N., D.R. McCabe and T. Maber, 1997. The
damage (Rikans and Hornbrook,1997). The most
Analysis of Free Radicals, their Reaction Products
prominent ROS are the superoxide anion (O •–), hydrogen
and Antioxidants. In: Baskin, S.I. and H. Salem
peroxide (H O ), and the hydroxyl ion (OH•) (Turner and
(Eds.), Oxidants, Antioxidants and Free Radicals.
Lysiak, 2008). Cells also have intrinsic antioxidant
Chap. 2, Taylor and Francis, Washington, DC.
systems that counter ROS accumulation. These include
Adelusi, S.A. and L.A. Salako, 1982. Tissue and blood
enzymes such as catalase, glutathione peroxidases, and
concentration of chloroquine following chronic
superoxide dismutase, and nonenzymatic antioxidants,
administration in the rat. J. Pharm. Pharmacol., 34:
such as vitamins E, C, beta carotene, ubiquinone, lipotic
acid, and urate (Nordberg and Arner, 2001; Giordano,
Ajani, E.O., P.D. Shallie, B.O. Adegbesan, B.A. Salau
and M. Adesanya, 2008. Protective effect of garcinia
In a normal liver, the level of ROS is low, and
kola (kolaviron) extract on predisposition of rats to
antioxidant defenses are adequate to protect the liver from
cardiovascular diseases following separate
oxidative damage (Fernandez et al., 1997). Nevertheless,
administration of amodiaquine and artesunate. Afr. J.
under several situations, the rate of generation of ROS
exceeds that of their removal and oxidative stress occurs
Amaravadi, R.K., D. Yu, J.J. Lum, T. Bui,
(Giordano, 2005; Di Giulio et al., 1995; Halliwell and
M.A. Christophorou, G.I. Evan, A. Thomas-
Gutteridge, 1999; Livingstone, 2001). However, more
Tikhonenko and C.B. Thompson, 2007. Autophagy
severe oxidative stress can cause cell death and even
inhibition enhances therapy-induced apoptosis in a
moderate oxidation can trigger apoptosis, while more
Myc-induced model of lymphoma. J. Clin. Invest.,
intense stresses may cause necrosis (Lennon et al., 1991).
However, under the severe levels of oxidative stress that
Balla, J., H.S. Jacob, G. Balla, K. Nath, J.W. Eaton and
cause necrosis, the damage causes ATP depletion,
G.M. Vercellotti, 1993. Enothelial-cell heme uptake
preventing controlled apoptotic death and causing the cell
from heme proteins: induction of sensitization and
to simply fall apart (Lelli et al., 1998; Lee et al., 1999).
desensitization to oxidant damage. Proc. Natl. Acad.
In this study, chloroquine may have acted directly
through generation of high levels of free heme or ROS on
Balla, G., H.S. Jacob, J.W. Eaton, J.D. Belcher and
the hepatocytes, affecting their cellular integrity and
G.M. Vercellotti, 1991. Hemin: A possible
causing defect in membrane permeability and cell volume
physiological mediator of low density lipoprotein
homeostasis. In cellular necrosis, the rate of progression
oxidation and endothelial injury. Arterioscler
depends on the severity of the environmental insults. The
greater the severity of the insults the more rapid the
Barennes, H., T. Balima-Koussoube', N. Nagot,
progression of neuronal injury (Ito et al., 2003). The
J.C. Charpentier and E. Pussard, 2006. Safety and
principle holds true for toxicological insult to the brain
efficacy of rectal compared with intramuscular
and other organs (Martins et al., 1978). Thus, it may be
quinine for the early treatment of moderately severe
inferred from this result that chronic oral administration
malaria in children: randomised clinical trial. Br.
of chloroquine is toxic to the liver in wistar rats.
Beri, R. and R. Chandra, 1993. Chemistry and biology of
CONCLUSION
heme: Effect of metal salts, organometals, andmetalloporphyrins on heme synthesis and catabolism,
Our study revealed that chronic oral administration of
with special reference to clinical implications and
chloroquine causes cytoplasmic vacuolation; nuclear
interactions with cytochrome P-450. Drug Metab.
enlargement and vesiculation of the hepatocytes. These
results have established the hepatotoxic potential of
Borges, L.P., R. Brandao, B. Godoi, C.W. Nogueira and
chronic oral administration of chloroquine in wistar rats.
G. Zeni, 2008. Oral administration of diphenyl
It is therefore recommended that the drug be prescribed
diselenide protects against cadmium-induced liver
with caution in patients with history of liver disease.
damage in rats. Chem. Biol. Interact., 171: 15-25. Br. J. Pharmacol. Toxicol., 2(3): 97-103, 2011
Bose, S. and S.P. Sinha, 1994. Modulation of ochratoxin-
Giordano, F.J., 2005. Oxygen, oxidative stress, hypoxia,
produced genotoxicity in mice by vitamin C. Food
and heart failure. J. Clin. Invest., 115(3): 500-508.
Green, D.R., 1998. Apoptotic pathways: The roads to
Boya, P., R.A. González-Polo, N. Casares, J.L. Perfettini,
P. Dessen, N. Larochette, D. Métivier, D. Meley, S.
Grunewald, S., U. Paasch, T.M. Said, R.K. Sharma,
Souquere, T. Yoshimori, G. Pierron, P. Codogno and
H.J. Glander and A. Agarwal, 2005. Caspase
G. Kroemer, 2005. Inhibition of macroautophagy
activation in human spermatozoa in response to
triggers apoptosis. Mol. Cell. Biol., 25(3):
physiological and pathological stimuli. Fertil Steril,
Cohen, J.J., 1993. Apoptosis. Immunol. Today, 14: 126-
Halliwell, B. and J.M.C. Gutteridge, 1999. Free Radicals
in Biology and Medicine. 3rd Edn., Oxford
Cook, G.C., 1994. Malaria in the liver. Postgrad Med. J.,
Hien, T.T. and N.J. White, 1993. Qinghaosu. Lancet, 341:
Debra, K.F. and N.L. Megan, 1999. Quinine-induced
hepatotoxicity. Ann. Pharmacother., 33: 32-34.
Hsu, C.Y., Y.P. Chan and J. Chang, 2007. Antioxidant
deGroot, P.Q., R.Q. Eiferink, M. Hollemans, M. Khand
activity of extract from Polygonum cuspidatum. Biol.
and J.M. Tager, 1981. Activation of B galactosidase
in cultured human skin fibroblast. Exp. Cell. Res.,
Ito, U., M. Sparts, J.R. Walker and I. Warzo, 2003.
Experimental Cerebral Ischemia in Magolian Gerbils
Dhalla, A.K., M.F. Hill and P.K. Singal, 1996. Role of
oxidative stress in transition of hypertrophy to heart
failure. J. Am. Coll. Cardiol., 28(2): 506-514.
Izunya, A.M., A.O. Nwaopara, A. Aigbiremolen
Di Giulio, R.T., W.H. Benson, B.M. Sanders and
M.A.C. Odike, G.A. Oaikhena and J.K. Bankole,
P.A. Van Veld, 1995. Biochemical Mechanisms:
2010. Histological effects of oral administration of
Metabolism, Adaptation and Toxicity. In: Rand, G.
artesunate on the liver in wistar rat. Res. J. Appl. Sci.
(Ed.), Fundamentals of Aquatic Toxicology. Effects,
Environmental Fate and Risk Assessment. Taylor and
Konat, G.W., 2003. H O -induced higher order chromatin
degradation: a novel mechanism of oxidative
Drury, R.A.B., E.A. Wallington and R. Cameron, 1967.
genotoxicity. J. Biosci., 28 :57-60.
Carleton's Histological Techniques. 4th Edn., Oxford
Kumar, S. and U. Bandyopadhyay, 2005. Free heme
toxicity and its detoxification systems in human.
Ducharme, J. and R. Farinotti, 1996. Clinical
pharmacokinetics and metabolism of chloroquine.
Larrey, D., A. Castot, D. Pessayre, P. Merigot,
Focus on recent advancements. Clin. Pharm., 31:
J.P. Machaye khy, G. Feldmann, A. Lenoir,
B. Rueff and J.P. Benha mou, 1986. Amodiaquine
Dubois, E. L., 1978. Antimalarials in the management of
induced hepatitis: A report of seven cases. Ann. Int.
discoid and systemic lupus erythematorus. Semin
Lee, Y.J. and E. Shacter, 1999. Oxidative stress inhibits
El-Sharaky, A.S., A.A. Newairy, M.M. Badreldeen,
apoptosis in human lymphoma cells. J. Biol. Chem.,
S.M. Eweda and S.A. Sheweita, 2007. Protective
274(28): 19792-19798, doi: 10.1074/jbc.274.28.
role of selenium against renal toxicity induced by
cadmium in rats. Toxicology, 235(3): 185-193
Lelli, J.L., L.L. Becks, M.I. Dabrowska and
Fan, C., W. Wang, B. Zhao, S. Zhang and J. Miao, 2006.
D.B. Hinshaw, 1998. ATP converts necrosis to
Chloroquine inhibits cell growth and induces cell
apoptosis in oxidant-injured endothelial cells. Free
death in A549 lung cancer cells. Bioorg. Med.
10.1016/S0891-5849(98)00107-5. PMID: 9801070.
Farber, J.L., K.R. Chein and S. Mittnacht, 1981. The
Lennon, S.V., S.J. Martin and T.G. Cotter, 1991. Dose-
pathogenesis of Irreversible cell injury in ischemia.
dependent induction of apoptosis in human tumour
cell lines by widely diverging stimuli. Cell Prolif.,
Fernandez-Checa, J.C., N. Kaplowitz, C. Garcia-Ruiz,
24(2): 203-214. doi: 10.1111/j.1365-2184.tb01150.x.
A. Colell, M. Miranda, M. Mari, E. Ardite and
A. Morales, 1997. GSH transport in mitochondria:
Livingstone, D.R., 2001. Contaminated-stimulated
Defense against TNFinduced oxidative stress and
reactive oxygen species production and oxidative
alcohol-induced defect. Am. J. Physiol. Gastrointest
damage in aquatic organisms. Mar. Poll. Bull., 42:
Br. J. Pharmacol. Toxicol., 2(3): 97-103, 2011
Lockwood, T.D., 2000. Redox control of protein
Oforah, E., B.J. Idang and N. Kalu, 2004. Chronic
degradation. Antioxid. Redox Signal., 2: 851-878.
chloroquine administration causes low circulating
Maclean, K.H., F.C. Dorsey, J.L. Cleveland and
plasma testosterone and low luteinizing hormone
M.B. Kastan, 2008 Targeting lysosomal degradation
associated with testicular lesion in rat. Acta Pharm.
induces p53-dependent cell death and prevents cancer
in mouse models of lymphomagenesis. J. Clin.
S.K. Akindele, 1997. A study of the hepatotoxicity of
Makker, K., A. Agarwal and R. Sharma, 2009. Oxidative
chloroquine (SN-7618) in mice. Nig. Qt. J. Hosp.
stress and male infertility. Indian J. Med. Res., 129:
Olanrewaju, W.I. and A.W.B.R. Johnson, 2001.
Martins, L.J., N.A. Al-Abdulla, J.R. Kirsh, F.E. Sieber
Chloroquine-resistance Plasmodium falciparummalaria in Ilorin, Nigeria: Prevalence and risk factors
and C. Portera-Cailliau, 1978. Neurodegeneration in
for treatment failure. Afr. J. Med. Sci., 30: 165-169.
excitotoxicity, global cerebral ischaemia and target
Papa, S. and V.P. Skulachev, 1997. Reactive oxygen
Deprivation: A perspective on the contributions of
species, mitochondria, apoptosis and aging. Mol.
apoptosis and necrosis. Brain Res. Bull., 46(4):
Pero, O. and W.R.J. Taylor, 2002. Amodiaquine for the
McConkey, D.J. and S. Orrenius, 1991. In: Tomei, L.D.
treatment of uncomplicated Falciparum malaria.
and F.O. Cope (Eds.), Apoptosis: The Molecular
Basis of Cell Death. Cold Spring Harbor Laboratory
Pollman, M.J., T. Yamada, M. Horiuchi and
G.H. Gibbons, 1996. Vasoactive substances regulate
Mitscher, L.A., 2005. Bacterial topoisomerase inhibitors:
vascular smooth muscle cell apoptosis. Circ. Res.,
Quinolone and pyridone antibacterial agents. Chem.
Rahman, I., 2003. Oxidative stress, chromatin remodeling
Murdoch, C.E., M. Zhang, A.C. Cave and A.M. Shah,
and gene transcription in inflammation and chronic
2006. NADPH oxidase-dependent redox signalling in
lung diseases. J. Biochem. Mol. Biol., 36: 95-109.
cardiac hypertrophy, remodelling and failure.
Razinger, J., M. Dermastia, J.D. Koce and A. Zrimec,
2008. Oxidative stress in duckweed (Lemna minor
Neftel, K.A., W. Woodtly, M. Schmid, P.G. Frick and
L.) caused by short-term cadmium exposure.
J. Fehr, 1986. Amodiaquine induced agranulocytosis
and liver damage. Br. Med. J., 292: 721-723.
Reisinger, E.C., R.D. Horstmann and M. Dietrich, 1989.
Ngaha, E.O., 1982. Some biochemical changes in the rat
Tolerance of mefloquine alone and in combination
during repeated chloroquine administration. Toxicol.
with sulfadoxine-pyrimethamine in the prophylaxis
of malaria. Trans. R. Soc. Trop. Med. Hyg., 83:
Ngokere, A.A., T.C. Ngokere and A.P. Ikwudinma, 2004.
Acute study of histomorphological and biochemical
Rikans, L.E. and K.R. Hornbrook, 1997. Lipid
changes caused by artesunate in visceral organs of
peroxidation, antioxidant protection and aging.
the Rabbit. J. Exp. Clin. Anat., 3(2): 11-16(s).
Biochem. Biophys. Acta, 1362: 116-127.
Nordberg, J. and E.S. Arner, 2001. Reactive oxygen
Sakkas, D., E. Mariethoz, G. Manicardi, D. Bizzaro,
P.G. Bianchi and U. Bianchi, 1999. Origin of DNA
species, antioxidants, and the mammalian thioredoxin
damage in ejaculated human spermatozoa. Rev.
system. Free Radic. Biol. Med., 31: 1287-1312.
Nosten, F. and P. Brasseur, 2002. Combination therapy
Sawyer, D.B., D.A. Siwik, L. Xiao, D.R. Pimentel,
for malaria: The way forward? Drug, 62(9):
K. Singh and W.S. Colucci, 2002. Role of oxidative
stress in myocardial hypertrophy and failure. J. Mol.
Nwanjo, H., I. Iroagba, I. Nnatuanya and N. Eze, 2007.
Antifertility activity of dihydroartemisinin in male
Schulte-Hermann, R., W. Bursch, B. Marian and
Albino rats. Internet J. Endocrinol., 4(1), ISSN:
B. Grasl-Kraupp, 1999. Active cell death (apoptosis)
and cellular proliferation as indicators of exposure to
Nwanjo, H. and G. Oze, 2007. Acute hepatotocixity
carcinogens. IARC Scientific Publications (Lyon),
following administration of artesunate in guinea pigs.
Internet J. Toxicol., 4(1), ISSN: 1559-3916
Shacka, J.J., B.J. Klocke, M. Shibata, Y. Uchiyama,
Obi, E., O.E. Orisakwe, L.A. Asomugha and
G. Datta, R.E. Schmidt and K. A. Roth, 2006.
O.O. Udemezue, 2004. The hepatotoxic effect of
Bafilomycin A1 inhibits chloroquine-induced death
halofantrine in guinea pigs. Indian J. Pharm., 36(5):
of cerebellar granule neurons. Mol. Pharmacol., 69:
Br. J. Pharmacol. Toxicol., 2(3): 97-103, 2011
Sharma, A. and M.C. Mishra, 1999. Inhibition of a
Vaux, D.L., G. Haecker and A. Strasser, 1994. An
protein tyrosine kinase activity in Plasmodium
evolutionary perspective on apoptosis. Cell, 76:
falciparum by chloroquine. Indian J. Biochem.
Wagener, F.A., H.D. Volk, D. Willis, N.G. Abraham,
Shen, H.M., J. Dai, S.E. Chia, A. Lim and C.N. Ong,
M.P. Soares, G.J. Adema and C.G. Figdor, 2003.
2002. Detection of apoptotic alterations in sperm in
Different faces of hemeoxygenase system in
subfertile patients and their correlations with spermquality. Hum. Reprod., 17: 1266-1273.
inflammation. Pharmacol. Rev., 55: 551-571.
Sinha, H.A.P. and R.S. Swerdloff, 1999. Hormonal and
Wernsdorfer, W.H. and I. McGregor, 1988. Malaria:
genetic control of germ cell apoptosis in the testis.
Principles and Practice of Malariology. Churchill
Stadtman, E.R., and R.L. Levine, 2003. Free
World Health Organization, 1990. Practical chemotherapy
radicalmediated oxidation of free amino acids and
of malaria. Report of a WHO scientific group,
amino acid residues in proteins. Amino Acids, 25:
Geneva. World Health Organization Technical
Thompson, C.B., 1995. Apoptosis in the pathogenesis and
Wyllie, A.H., 1980. Glucocorticoid-induced thymocyte
treatment of disease. Science, 267: 1456-1462.
Turner, T. and J.J. Lysiak, 2008. Oxidative stress: A
common factor in testicular dysfunction. J. Androl.,
endonuclease activation. Nature, 284: 555-556.
29(5): 488-498. doi: 10.2164/jandrol.108.005132.
Hydroxycut Review By Elizabeth Myers, RD Hydroxycut is currently of the most familiar weight-loss supplements on the market. It is heavily advertised on television and sold in 70 countries worldwide. Hydroxycut claims to cause fast weight loss, increased energy, and a controlled appetite. The supplement costs $44.99/bottle for 150 capsules, which is quite expensive for a weight loss sup
Chapter 2. Empiric Therapy: CNS Infections Encephalitis Usual Pathogens IV-to-PO Switch Acyclovir 10 mg/kg (IV) q8h x 7 days, then if able to take oralmedications, complete 14-21 days of total therapy withacyclovir 400 mg (PO) 5x/day or valacyclovir 1 gm (PO) q8h or famciclovir 500 mg (PO) q8hUsual PathogensCalifornia encephalitis (CE), Western equine encephalitis (WEE), Venezuelan e
 Br. J. Pharmacol. Toxicol., 2(3): 97-103, 2011
inactivated in the liver and is therefore susceptible to thetoxicity from these agents. Certain medicinal agents whentaken in overdoses and sometimes even when introducedwithin therapeutic ranges may injure the liver.
Reports regarding the effects of chronic oral
administration of chloroquine on the histology of the liverare scanty in existing literatures. There is however areport which showed that chloroquine treatment for 12weeks in mice causes cytolysis in hepatocytes (Okonkwoet al., 1997).
This study was considered important since
rheumatoid arthritis and malaria are common ailments inthe tropics and the need to avoid the risk of hepatitisresulting from prolonged oral administration ofchloroquine. In view of this, the present study wascarried out to investigate the effect of chronic oral
Plate 1: (Control Group): Control section of the liver showing
administration of chloroquine on the histology of the liver
MATERIALS AND METHODS
Br. J. Pharmacol. Toxicol., 2(3): 97-103, 2011
inactivated in the liver and is therefore susceptible to thetoxicity from these agents. Certain medicinal agents whentaken in overdoses and sometimes even when introducedwithin therapeutic ranges may injure the liver.
Reports regarding the effects of chronic oral
administration of chloroquine on the histology of the liverare scanty in existing literatures. There is however areport which showed that chloroquine treatment for 12weeks in mice causes cytolysis in hepatocytes (Okonkwoet al., 1997).
This study was considered important since
rheumatoid arthritis and malaria are common ailments inthe tropics and the need to avoid the risk of hepatitisresulting from prolonged oral administration ofchloroquine. In view of this, the present study wascarried out to investigate the effect of chronic oral
Plate 1: (Control Group): Control section of the liver showing
administration of chloroquine on the histology of the liver
MATERIALS AND METHODS